Генофонд.рф / Народы и регионы / Европа / Геногеография гаплогрупп Y-хромосомы в Европе
Геногеография гаплогрупп Y-хромосомы в Европе
17.11.2015
Скачать страницу в PDF
 Структура генофонда Европы по Y-хромосоме: «принцип паззлов».
Анализ генофонда Европы начнем с рассмотрения геногеографии отдельных гаплогрупп. Отчасти это является продолжением предыдущего раздела, в котором кратко суммировалось то, что было известно о распространении гаплогрупп Y-хромосомы до нашего исследования, но теперь мы рассмотрим современные, подробные карты распространения всех основных европейских гаплогрупп и тех их субвариантов, для которых уже известны частоты в ряде популяций Европы.
В нашей базе данных Y-base содержится почти вся опубликованная информация о частотах гаплогрупп Y-хромосомы в мире (и в Европе в частности), но далеко не вся эта информация пригодна для изучения географического распространения гаплогрупп. Как и при создании других геногеографических атласов, для создания карт распространения Y-хромосомы в Европе данные были тщательно отобраны и создан массив данных для карт Y-хромосомы в Европе. Рассмотрим, по каким критериям этот массив данных формировался.
ОРГАНИЗАЦИЯ ДАННЫХ
МАССИВ ДАННЫХ ДЛЯ КАРТ: ПОПУЛЯЦИИ.
Для надежного определения частоты однородительских генетических маркеров (Y-хромосомы и мтДНК) требуется объем выборки в два раза больший, чем для аутосомных маркеров – поэтому нижняя граница выборки должна быть не менее 70-100 образцов. Маленькие выборки или объединялись с географически соседними выборками того же народа, или (если объединение было невозможно) не включались в анализ. В результате средний объем выборки в массиве данных для карт составляет N=136 образцов. Но делались и исключения из правила. По некоторым народам или же региональным группам в пределах народа, которые для полноты картины хотелось обязательно представить на карте, были доступны лишь небольшие выборки. Тогда, скрепя сердце, приходилось включать в массив для карт и выборки меньшего размера. Но такие исключения делались редко — из 251 картографированных популяций лишь десятая часть (29 популяций) имеют объем выборки ниже N=50, а подавляющее большинство (192 выборки) имеют объем N=70 и выше.
Если для какого-либо народа были данные по его региональным популяциям, то для картографирования не использовались выборки этого народа, для которых географическая привязка неизвестна (если авторы при публикации указали лишь принадлежность к народу).
В итоге массив для карт включил данные по 251 популяциям (выборкам), и суммарное число образцов во всех выборках составило 34 294 образцов. Стоит отметить, что значительная часть этих данных получена благодаря исследованиям нашего коллектива под руководством автора этих строк: 45 популяций и 4 880 образцов. Это наши данные по русским популяциям [Balanovsky et al., 2008], украинским, белорусским и некоторым дополнительным русским популяциям [Kushniarevich et al., 2015], популяциям Северного Кавказа [Balanovsky et al., 2011], а также наши еще неопубликованные данные по популяциям крымских и казанских татар, мокши и эрзи. (Данные по частотам гаплогрупп из трех опубликованных статей приведены в таблицах 2.1, 2.2, 2.3.) Получается, что из всего массива данных по изменчивости Y-хромосомы в Европе каждая пятая популяция и каждый седьмой образец изучены нашим коллективом ( на рис 2.1. «наши» популяции показаны синими кружками), хотя исследованиями в этой области занималось и занимается несколько десятков лабораторий по всей Европе. Если добавить, что Европа — самый изученный регион мира, то это отчасти объясняет, почему именно нашим коллективом предпринят обобщающий анализ генофонда Европы. Для построения карт использованы также данные Y-base не только по европейским популяциям, но и по популяциям близлежащих регионов, входящих в картографируемый ареал.
Кроме 251 популяций Европы, изученных по панелям гаплогрупп, охватывающих весь их спектр, еще около 200 популяций были изучены по субвариантам только одной какой-то гаплогруппы. В основном это статьи [Myres et al., 2010; Underhill et al., 2010, 2014], посвященные гаплогруппам R1b и R1a, и ряд статей по гаплогруппам N, E, и J. Как правило, выборки, использованные в этих статьях для углубленного генотипирования отдельных гаплогрупп, раньше уже изучались по всему спектру гаплогрупп. Рис 2.1. дает представление о плотности покрытия Европы и смежных регионов популяциями, изученными не по одной гаплогруппе, а по всему их спектру с обычным уровнем филогенетического разрешения.
МАССИВ ДАННЫХ ДЛЯ КАРТ: ГАПЛОГРУППЫ.
Это понятие «обычного филогенетического уровня» изученности гаплогрупп очень условно, и уровень этот растет с каждым годом. Если в 2000 году число гаплогрупп в обычной статье не дотягивало до десятка, то в 2010е годы большинство статей используют панели из 40-70 гаплогрупп. Конечно, начиная примерно с 2013 года, число известных гаплогрупп, обнаруженных благодаря полному секвенированию, исчисляется уже тысячами. Но обнаружение их — только первый шаг, который требует множество следующих шагов и усилий по анализу большого массива самых разных популяций на наличие новых гаплогрупп. И пока не проведено определение их частот в широком спектре популяций, значение этих новооткрытых гаплогрупп для исследований генофонда невелико.
Объединяя в одной таблице данные из десятков статей разных лет, неизбежно оказывается, что некоторые гаплогруппы изучены в очень большом числе популяций, а некоторые – лишь для немногих популяций. К счастью, для картографирования это не представляет большой проблемы (уж в двух-трех десятках популяций изучены почти все отобранные для картографирования гаплогруппы, а такого числа опорных точек обычно достаточно для выявления основных трендов). Но все же при рассмотрении карт стоит иметь в виду, что подробность исходных данных для разных гаплогрупп неодинакова.
Для картографирования отбирались гаплогруппы, составляющие заметную часть генофонда хотя бы в некоторых популяциях Европы и при этом изученные, как только что было сказано, в нескольких десятках популяций. Этим критериям удовлетворили 40 гаплогрупп: E1b1a1-DYS271, E1b1b1a1b-L142.1, E1b1b1a1c-V22, E1b1b1b1-M81, E1b1b1c-M123, G2a1-P16, G2a3a-M406, G2a3b1-P303, I1-L118, I2a-P37.2, I2b-L35, J1-L255, J1c3-P58, J2-L228, J2a-L152, J2a3b-M67, J2a3h-L207.1, J2b-L282, L-M11, N1b-P43, N1c-M46, O3-M122, Q-M242, R1a1-L120, R1a1a1g-M458, R1a-M558, R1a-Z284, R1a-Z93, R1b1a1-M478, R1b1a2-L265, R1b-L10, R1b1a2a1a1a-M405, R1b1a2a1a1a1-M467, R1b1a2a1a1b-P312, R1b1a2a1a1b2a-M153, R1b1a2a1a1b2b1-M167, R1b1a2a1a1b4b-M222, R1b1c-V88, R2a-L266, T-L206.
Таблица 2.1. Наши данные о частотах гаплогрупп Y-хромосомы в русских популяциях
(опубликованы в [Balanovsky et al., 2008]).
 Примечания: 1 К гаплогруппе F* относятся образцы с производным аллелем маркера М89, но с предковыми аллелями по маркерам M9, M201, M170, 12f2. 2 К гаплогруппе G1* относятся образцы с производным аллелем маркера M285, но с предковым по маркеру P20. 3 К гаплогруппе K* относятся образцы с производным аллелем по маркеру M9, но с предковыми по маркерам M70, 92R7, M231. 4 К гаплогруппе N* относятся образцы с производным аллелем по маркеру M231, и с предковыми по маркерам TAT, P43, M128.
Таблица 2.2. Наши данные о частотах гаплогрупп Y-хромосомы в популяциях балто-славянских народов (опубликованы в [Kushniarevich et al., 2015]).
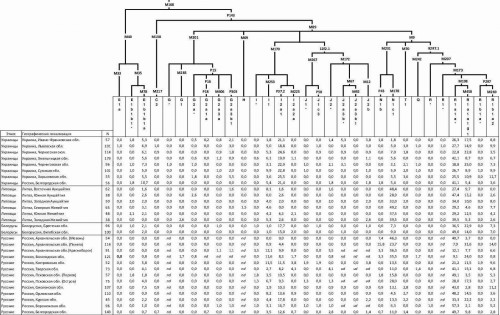
Таблица 2.3. Наши данные о частотах гаплогрупп Y-хромосомы в популяциях Кавказа
(опубликованы в [Balanovsky et al., 2011]).
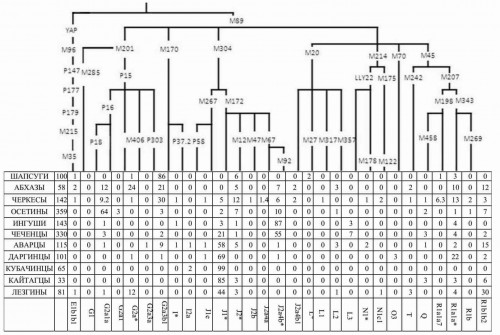
 Рис. 2.1. Географическое положение популяций Европы и смежных регионов, изученных по полиморфизму Y-хромосомы. Синими кружочками показаны наши собственные данные, зелеными — данные из литературы.
ГАПЛОГРУППА R1a И ЕЕ ВАРИАНТЫ
ГАПЛОГРУППА R1a (без подразделения на субветви).
Как указывалось в обзоре предшествовавших работ, гаплогруппа R1a является одной из двух наиболее частых гаплогрупп Европы. Более того, она является и наиболее широко распространенной гаплогруппой в Евразии в целом. Для ее генотипирования используются разные маркеры – M198, M17, SRY1542, филогенетическая разница между которыми почти отсутствует, и известно множество других маркеров, определяющих эту гаплогруппу, поэтому для ее обозначения может использоваться любой из этих них. В массиве данных для карт она фигурирует как R1a-L120.
 Y-хромосомы в Европе.") Рис. 2.2. География гаплогруппы R-L120 (R1a) Y-хромосомы в Европе.
На рис. 2.2 можно видеть четкий тренд географического распространения R1a-L120: хотя она распространена по всей Европе, но обширная зона максимальных частот находится в Восточной Европе, причем в этих популяциях она составляет почти половину генофонда. На востоке зону максимума ограничивает Волга, причем на всем ее протяжении – от Верхней Волги до низовий. На юге зона максимума доходит почти до Черного моря, на северо-западе – до Балтийского моря, а на юго-западе ограничивается Карпатами и Альпами. Но наиболее любопытны границы этой области на западе: хотя никаких географических барьеров там нет, зона высоких частот R1a все равно довольно четко ограничена, и падение частоты на каких-то двухстах-трехстах километрах достигает почти 30% (от примерно 35-45% у поляков и сорбов до 10-15% у немцев). Анализу этой «польско-немецкой генетической границы» были посвящены несколько статей [Ploski et al., 2002; Kayser et al., 2005; Rebala et al., 2013][1].
Я не упомянул еще границу зоны максимальных частот R1a на севере – хотя перепад частот там не так выражен, как на западе (не от красных к зеленым, а только от красных к желтым тонам, то есть процентов на пятнадцать), но северная граница тоже выявляется на карте вполне рельефно. В данном случае она проходит внутри ареала одного народа – русского – маркируя отличия генофонда Русского Севера от остальных русских популяций. Эти желтые тона умеренных частот занимают не только Русский Север, но и все пространство Восточной Европы между Волгой и Уралом, и лишь за Уральским хребтом частоты R1a падают до столь же низких значений (зеленые тона на карте), что и в Западной Европе, Скандинавии, Балканах и Передней Азии.
СУБВАРИАНТЫ ГАПЛОГРУППЫ R1a
В пределах такого огромного ареала гаплогруппы R1a-L120, несомненно, существует почти столь же огромное число субвариантов этой гаплогруппы, причем многие из них могут иметь четкие и сравнительно небольшие географические ареалы. Но в течение долгого времени субтипы в пределах R1a были неизвестны. Пока, благодаря работам лаборатории Peter Underhill в Стэнфордском университете не были открыты сначала первые два, а потом и еще несколько субвариантов. Конечно, в настоящее время число известных субвариантов R1a исчисляется сотнями: каждый образец, для которого проведено полное секвенирование всего генома или Y-хромосомы, образует свою ветвь на филогенетическом древе. Но пока хотя бы для некоторых из этих вариантов не проведен скрининг их распространенности в популяциях и не определены частоты их встречаемости в разных популяциях Европы, построение геногеографических карт невозможно. Сейчас об ареалах распространения этой субветвей можно рассуждать лишь гадательно, основываясь на порой зыбких сведениях о местах происхождения тех единичных образцов, по которым каждая из этих сотен ветвей была открыта. Интерес к этой гаплогруппе столь велик, что, конечно, такой популяционный скрининг будет проведен разными лабораториями мира (в том числе и нашим коллективом) в самые ближайшие годы, и тогда геногеография субвариантов R1a станет на порядок подробнее. Но на данный момент мы ограничены теми маркерами, по которым проведен массовый популяционный скрининг в статьях [Underhil et al., 2010; 2014] и в работах нашего коллектива: это три основных маркера европейской ветви R1a и маркер азиатской ветви.
АЗИАТСКАЯ ВЕТВЬ R1A-Z93.
Надо отметить, что деление на европейскую и азиатскую ветви проявляется на филогенетическом древе R1a очень четко [Karmin et al., 2015]. Маркером азиатской ветви является Z93 (и его синонимы), и в Европе эта ветвь почти отсутствует. В доказательство этого приведем карту ее распространения (рис. 2.3.) на которой видны только низкие частоты (1-3%) и только в очень немногих популяциях Восточной Европы, и перейдем к рассмотрению европейских субвариантов.
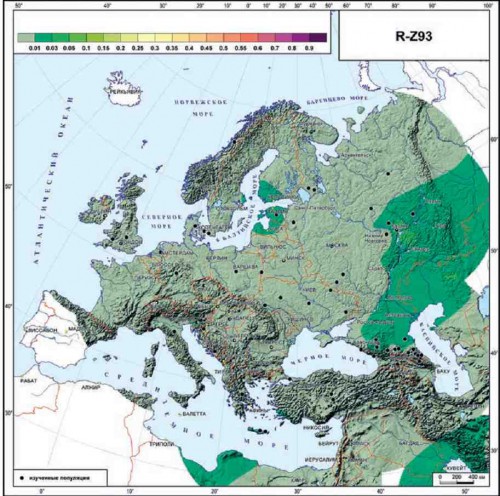
Рис. 2.3. География гаплогруппы R-Z93 Y-хромосомы в Европе.
ЕВРОПЕЙСКАЯ ВЕТВЬ R1A-М458.
Первый из них, R-М458, выделяет центрально-европейский субвариант в общем массиве гаплогруппы R1a, занимающей половину Европы. Поэтому после открытия этого маркера, мы специально генотипировали его во многих славянских и северокавказских популяциях, что позволило впервые построить подробную карту его распространения с учетом Восточной Европы и Кавказа (рис. 2.4).
 Рис. 2.4. География гаплогруппы R-M458 Y-хромосомы в Европе.
Карта демонстрирует наиболее высокие частоты этого маркера у западных славян (поляков и чехов), где он составляет четверть генофонда, и несколько сниженные частоты в окружающих популяциях. Распространение этого маркера на запад ограничено прилежащими популяциями немцев (вероятно, ассимилировавших западнославянские группы, ранее населявшие эти территории). Зато на восток зоны высоких частот гаплогруппы R-M458 распространяется далеко, захватывая весь ареал восточных славян. Характерно, что эта восточная экспансия более выражена в лесостепной зоне (у украинцев), чем в лесной (белорусы и русские). Еще более любопытным представляется ограниченность экспансии на юг: у южных славян и неславянских народов Балкан (венгры, румыны) частота этой гаплогруппы мала, причем прослеживается соответствие генетической «границы» высоких частот R-M458 и географической границы Карпат. К северу от этого горного хребта расположена зона максимальных частот гаплогруппы, а к югу от Карпат частота резко снижается. По-видимому, Карпаты выступили в роле географического барьера для потока генов, препятствуя распространению этой гаплогруппы на Балканы. Отметим, что в отличие от R1a в целом, встречающейся хотя бы с низкими частотами в любом регионе Европы, R-M458 распространена только по всей Восточной Европе, соседних территориях Западной Европы и на Балканах, но практически полностью отсутствует на большей части Западной Европы – Пиренейском полуострове, Франции, Британских островах, значительной части Аппенинского и Скандинавского полуостровов, а также отсутствует за пределами Европы – и в Передней Азии, и в Сибири.
ЕВРОПЕЙСКАЯ ВЕТВЬ R1A-М558.
Второй европейский субвариант R1a обозначается по названию маркера R-M558 (рис. 2.5.) Филогенетически он братский (для мтДНК в таких случаях принято говорить «сестринский», но тут речь идет о Y-хромосоме) для R-M458. Оба этих маркера распространены сходным образом: R-M558 тоже распространен преимущественно в Восточной Европе, тоже реже встречается на Балканах, заходит с низкими частотами в соседние области Западной Европы, и так же, как и R-M458, практически отсутствует за пределами Европы. Ареалы этих субтипов перекрываются и по зоне их максимальных частот у западных и восточных славян. Основное различие между паттернами распространения R-M458 и R-M558 состоит только в большей приуроченности к западной или восточной части их общего ареала.
 Рис. 2.5. География гаплогруппы R-M558 Y-хромосомы в Европе.
R-M458 имеет склонность к западу ареала. У западных славян встречается в среднем с частотой 25% (т.е. четверть генофонда), а у восточных славян в среднем встречается с частотой 15%, хотя и повышается до 23% у западных белорусов и у некоторых украинских популяций (но не западных, а центральных).
R-M558 имеет склонность к востоку их общего ареала. Он встречается с частотой выше 30% в обеих изученных по этому маркеру популяциях южных русских; составляет 20-25% у белорусов, поляков, словаков, западных украинцев; снижается ниже (или значительно ниже) 20% у центральных русских, чехов, центральных и восточных украинцев и чехов. Впрочем, R-M558 изучен в значительно меньшем числе популяций, чем R-M458, и поэтому карта его распространения может еще значительно уточниться в будущем. Мы видим, что даже это основное различие между паттернами распространения R-M458 и R-M458 выражается в небольших различиях частот и имеет довольно сложную географию.
Этот пример ясно показывает, что повышение филогенетического разрешения далеко не всегда приводит к повышению географического разрешения: хотя в пределах европейской ветви R1a и удалось выделить два субтипа, но их географическое распространение очень сходно. По всей видимости, они возникли в родственных популяциях и распространялись совместно, в ходе одних и тех же миграций. Или же, возникнув даже в разных частях ареала популяций, интенсивно обменивавшихся миграциями друг с другом, они распространились в пределах всего этого ареала. Конечно, повышать филогенетическое разрешение необходимо, и, как правило, выделяемые субтипы имеют более четкую географическую привязку, чем ветвь в целом; субтипы следующего уровня имеют еще более узкую зону распространения в пределах зон субтипов предыдущего уровня и так далее. Но эта географическая четкость зависит не только и даже не столько от уровня филогенетического разрешения, сколько от структуры генофонда, от исторически заданного соотношения изоляции и метисации, то есть, в терминах популяционной генетики, от баланса дрейфа генов и миграций.
Если этот баланс сдвинут в сторону изоляции (как на Кавказе или в Сибири), то частоты гаплогрупп будут резко различаться между популяциями, гаплогруппы будут иметь географически узкие ареалы, мало перекрывающиеся друг с другом, и гаплогруппы будут достигать очень высоких частот в «своих» популяциях. Если же баланс сдвинут в сторону миграций, то частоты будут различаться не столь драматически, ареалы будут довольно широкими и перекрывающимися друг с другом, а субгаплогруппы достигать не слишком высоких частот, потому что остальная часть генофонда будет представлена другими гаплогруппами, ареалы которых заходят на эту же территорию. Все эти черты хорошо видны на примере R-M458 и R-M558, потому что сдвиг баланса в сторону не изоляции, а миграций особенно характерен именно для Европы.
ЕВРОПЕЙСКАЯ ВЕТВЬ R1A-Z284.
Зато еще один субвариант R1a – гаплогруппа R-Z284 (рис. 2.6.) имеет совершенно другой паттерн, который можно назвать «скандинавским» или «североевропейским». По имеющимся на настоящий момент данным [Underhill et al., 2014] его частота составляет 20% в Норвегии, 7% в Дании, и по 3% в Англии и Швеции, а за пределами Северной Европы R-Z284 встречена лишь у единичных образцов. Эти частоты наверняка скорректируются в ходе изучения новых популяций и увеличения объемов выборок уже изученных народов, но общая приуроченность к популяциям Северной Европы несомненна уже сейчас.
 Рис. 2.6. География гаплогруппы R-Z284 Y-хромосомы в Европе.
ГАПЛОГРУППА R1b И ЕЕ ВАРИАНТЫ
ГАПЛОГРУППА R1b (В ЦЕЛОМ)
Гаплогруппа R1b, «братская» по отношению к гаплогруппе R1a и вторая среди наиболее частых в Европе, имеет тоже широкий ареал (Рис. 2.8), однако основная зона её максимальных частот приходится на Западную Европу. Важным вкладом в изучение генофонда Европы стало открытие новых информативных SNP маркеров в пределах одной из ветвей гаплогруппы R1b — гаплогруппы R-М269 (на картах обозначена с помощью филогенетически близкого маркера как R-L265). Это масштабное исследование выполнено большим международным коллективом с участием автора и опубликовано в [Myres et al., 2010].
Карта распространения гаплогруппы R1b в целом (R-L10, рис. 2.8.) ярко показывает, что это основной компонент, составляющий более половины Y-хромосомного генофонда Западной Европы. Лишь население южной части Аппенинского полуострова несет сниженные частоты этой гаплогруппы, поскольку у них преобладают средиземноморские гаплогруппы, да еще Скандинавия по географии гаплогрупп относится не к Западной Европе, а формирует самостоятельный северный домен, в котором преобладает иная гаплогруппа — I1.
Гаплогруппа R1b даже в большей степени свойственна Западной Европе, чем R1a свойственна Восточной, поскольку R1b во многих популяциях (Британские острова, баски и ряд других популяций Пиренейского полуострова) составляет даже не половину, а более двух третей генофонда. В целом, тренд гаплогруппы R1b — максимальные частоты у басков и постепенное снижение частоты к востоку от Пиренейского полуострова — очень напоминает географию классического маркера Rh-d (резус-фактор). Всплеск частоты на востоке Европы отражает повышенную частоту R1b у некоторых популяций башкир, известную из работы [Лобов, 2009]. Впрочем, нашим коллективом сейчас завершается подробное исследование разнообразия Y-хромосомы в популяциях башкир (более 1000 образцов). В этом исследовании выявлено, что частота R1b велика только в немногих родовых группах, тогда как для большинства родовых групп башкир характерны другие гаплогруппы, а частоты R1b у них невелики. Поэтому всплеск R1b на востоке Европы отражает не общеевропейскую закономерность, а лишь локальные закономерности происхождения отдельных родовых групп степных популяций Евразии.
Хотя R1b достигает высоких частот в основном в Западной Европе, карта (рис. 2.8.) показывает, что она (как и R1a) распространена повсеместно, занимая весь картографированный ареал и уходя далеко за его пределы по Евразии. Считается, что R1b имеет переднеазиатское происхождение, и разные ее ветви распространились по различным уголкам Старого Света, но особенно «повезло» лишь той ветви, которая распространилась в Европе и достигла в ней столь значительных частот.
 Рис. 2.7. Филогенетическое дерево гаплогруппы R1b-L10.
На рисунке показаны только ветви, которые подробно рассматриваются с помощью геногеографических карт; структура схемы соответствует полному дереву гаплогруппы R1b-L10 по версии ISOGG на октябрь 2015 г.
 Y-хромосомы в Европе.") Рис. 2.8. География гаплогруппы R-L10 (R1b) Y-хромосомы в Европе.
СУБВАРИАНТЫ ГАПЛОГРУППЫ R1b
Карты на рис. 2.9, 2.10 и 2.11 показывают три основные ветви R1b (Рис. 2.7.), которые – со множеством оговорок – можно назвать африканской, степной евразийской и западноевропейской гаплогруппами. Действительно, когда мы смотрим на карту R1b в целом (рис. 2.8.), то видим ее наличие в том числе и в Северной Африке.
АФРИКАНСКАЯ ВЕТВЬ R-V88.
Карта R-V88 (рис. 2.9.) как раз и показывает частоты пусть и невысокие (1-6%, с повышением до 26% только в одной популяции), но зато по всей Северной Африке (от Марокко до Египта), и почти полное отсутствие этой гаплогруппы в Европе и в Азии. То есть «африканская» часть R1b – это почти исключительно ветвь R-V88. Своего максимума она достигает далеко за пределами картографируемого ареала – в Центральной Африке [Cruciani et al., 2010].
 Рис. 2.9. География гаплогруппы R-V88 Y-хромосомы в Европе.
СТЕПНАЯ ВЕТВЬ R-M478.
Карта следующей ветви – R-M478 – показывает ее приуроченность к степным популяциям Евразии (Рис. 2.10). Зона ее максимальных частот также находится по большей части за пределами картографируемого ареала – на пространствах от Урала до Алтая. Впрочем, как можно видеть на карте, отдельными «языками» эта гаплогруппа доходит и до Кавказа, оставаясь приуроченной к степным областям Южного Урала и Предкавказья, и составляя только 1-10%, редко в каких популяциях до 20% генофонда.
 Рис. 2.10. География гаплогруппы R-М478 Y-хромосомы в Европе.
 Рис. 2.11. География гаплогруппы R-L265 Y-хромосомы в Европе.
ЗАПАДНОЕВРОПЕЙСКАЯ ВЕТВЬ R-L265.
Третья из основных ветвей гаплогруппы R1b — гаплогруппа R-М269, или, как её здесь обозначаем на картах, R-L265 — составляет основную (по частоте и ареалу) часть R1b. Поэтому карта ее распространения (рис. 2.11) в целом вторит карте L10, лишь без африканской части ее ареала. Эта гаплогруппа подразделена на множество субгаплогрупп, многие из которых специфичны для той или иной территории в пределах ареала ветви. Остановимся на двух основных ветвях внутри R-L265: центральноевропейской гаплогруппе R-M405 и её пиренейско-британском «брате» R-P312 со всеми их «сыновними» ветвями (Рис. 2.12. — 2.17.).
 Рис. 2.12. География гаплогруппы R-M405 Y-хромосомы в Европе.
ЦЕНТРАЛЬНОЕВРОПЕЙСКАЯ ВЕТВЬ R-M405 В ЦЕЛОМ.
Гаплогруппа R-M405 (Рис. 2.12) встречается практически по всей Западной, Центральной и Северной Европе (за исключением южных областей Пиренейского и Аппенинского полуостровов, а также Балканского полуострова – на юге, Финляндии и Кольского полуострова – на севере), а также и в западной части Восточной Европы. Однако с максимальными частотами (18-36%) гаплогруппа R-M405 встречается преимущественно в населении Центральной Европы (на территории Германии, Швейцарии, Бельгии, Голландии, Дании и юге Британии). Датировки как возникновения, так и выделения внутренних субвариантов гаплогруппы R-M405 по данным www.yfull.com составляют около 5 тыс. лет назад (в интервале 4,4-5,3 тыс. лет назад), т.е. приходятся на бронзовый век. Похоже, что уже в это время происходили и миграционные процессы, приводящие к распространению гаплогруппы R-M405 в разные части Европы. Не исключено, что данные об гаплогруппе R-M405 указывают на один из эпизодов распространения носителей индо-европейских языков на территории Европы. На одно из направлений распространения носителей гаплогруппы R-M405, возможно, указывает география одной из её субветвей – гаплогруппы R-M467.
 Рис. 2.13. География гаплогруппы R-М467 Y-хромосомы в Европе.
«ГЕРМАНО-БРИТАНСКАЯ» ВЕТВЬ R-M467.
Скромное распространение гаплогруппы R-M467 (Рис. 2.13) — как по её частотам (от максимума 4% в восточной Германии через значения около 3% в северной и южной Германии, Голландии и на юге Британии до 0,5% во Франции), так и по географии (не самые обширные, по европейским масштабам, территории вокруг Северного моря) – тем не менее представляет интерес. География гаплогруппы R-M467 наводит на предположение о миграции её носителей с территории современной Германии вдоль Северного моря к берегам Британских островов. Датировки для гаплогруппы R-M467, представленные на сайте www.yfull.com, указывают на её возникновение около 4,7 тыс. лет назад (в интервале 4,1-5,4 тыс. лет назад), а время экспансии (разделения на субветви) спустя тысячу лет — около 3,4 тыс. лет назад (в интервале от 2,8-4,0 тыс. лет назад). Сопоставляя ареал гаплогруппы R-M467 на карте с указанными датировками возникновения и распространения, можно предположить две модели появления такой ситуации. Первая – когда миграционные процессы с территории центральных областей современной Германии времен бронзового века прошли вдоль берега Северного моря к Британским островам и оставили свой след в современном генофонде. Вторая – когда эта гаплогруппа могла быть распространена несколько шире на юг, запад и восток, но в результате прихода туда другого населения и без того невысокая частота гаплогруппы R-M467 снизилась на юге до величин, почти не улавливаемых при текущих объемах выборок.
ПИРЕНЕЙСКО-БРИТАНСКАЯ ВЕТВЬ R-P312 В ЦЕЛОМ.
Распространение гаплогруппы R-P312 (рис. 2.14) интересно тем, что во многом отражает общие особенности распределения в населении Европы как своей предковой гаплогруппы R-L265, так и всей гаплогруппы R1b в целом. Cамые высокие частоты гаплогруппы R-P312 на самом западе Европы (Пиренейский полуостров, территория западной Франции, Британские острова) с постепенным снижением частоты к востоку и локальный всплеск в южном Приуралье – это почти те же особенности, которые были описаны выше при характеристике ареала гаплогруппы R1b в целом. Однако следование общему тренду совсем не значит полное сходство. Так, обширная область красно-фиолетовых оттенков (частоты 50-75%) на территории большей части Западной Европы, которую наблюдали ранее на картах гаплогрупп R1b-L10 и R-L265 (Рис. 2.8. и Рис. 2.11.), на карте гаплогруппы R-P312 (Рис. 2.14) превращается в локальный всплеск частоты (до 75%) на севере Британских островов и область желто-красных оттенков (частота 30-50%) на территории Пиренейского полуострова и Франции. Южноуральский красно-фиолетовый всплеск общей частоты R1b на карте гаплогруппы R-P312 сокращается до небольшого «глазка» в точке для северных башкир, которую со всех сторон обтекают области низких частот (не более 3%).
Гаплогруппа R-Р312, таким образом, охватывает значительную часть как разнообразия всей гаплогруппы R-L265, так её ареала. Можно предполагать, что в таком случае внутри гаплогруппы R-P312 тоже существует ряд локальных субветвей с более узкими ареалами. И это предположение оправдывается: на сегодня известно пять больших ветвей, которые в свою очередь подразделяются на несколько десятков более глубоких. Однако остановимся только на трех ветвях R-P312, распространение которых в Европе хорошо изучено: гаплогруппа R-M167 и её субвариант R-M153, а также гаплогруппа R-M222 (Рис. 2.15-2.17.).
 Рис. 2.14. География гаплогруппы R-P312 Y-хромосомы в Европе.
«БАСКО-РОМАНСКАЯ» ГАПЛОГРУППА R-M167.
Такое название для гаплогруппы R-M167 условно: её география (рис. 2.15) охватывает население Пиренейского полуострова и Западной Франции, небольшую область на юге Германии и единичный «глазок» в Голландии, а еще область в Западном Причерноморье (румыны и болгары). Большинство ареала гаплогруппы, таким образом, населяют народы, говорящие на языках романской (испанцы, португальцы, французы и румыны) группы индоевропейской языковой семьи и баскской языковой семьи. Впрочем, южные немцы и болгары, у которых тоже встречена гаплогруппа R-M167, говорят на языках иных групп индоевропейской семьи.
Область максимальных частот гаплогруппы R-M167 (желтовато-зеленоватые тона на рис. 2.15, частота гаплогруппы от 6% до 25%) охватывает территорию Пиренейского полуострова и юго-западной Франции. Эта же область является единственной территорией распространения дочерней гаплогруппы R-M167 – субварианта R-M153 (Рис. 2.16).
 Рис. 2.15. География гаплогруппы R-M167 Y-хромосомы в Европе.
«БАСКСКАЯ» ВЕТВЬ R-M153.
Как можно проследить по карте (рис. 2.16), гаплогруппа R-M153 своей максимальной частоты (почти 16%) достигает у басков, в два раза реже встречается в населении испанских Пиренеев, и с очень низкими частотами (1-3%) охватывает оставшуюся часть своего ареала. Из-за такой географии гаплогруппу R-M153 можно условно назвать «баскской» и предположить, что она появилась и распространялась преимущественно в популяциях этого народа. Возраст возникновения этой гаплогруппы по данным www.yfull.com составляет около 2,5 тысяч лет (в пределах от 3500 до 1500 лет назад). Иными словами, эта гаплогруппа, вероятнее всего, появилась в популяциях Пиренейских гор на заре нашей эры и успела распространиться по обе стороны горного хребта (причем эффективнее – к югу от Пиренеев) в результате локальных миграций.
 Рис. 2.16. География гаплогруппы R-M153 Y-хромосомы в Европе.
 Рис. 2.17. География гаплогруппы R-M222 Y-хромосомы в Европе.
БРИТАНСКАЯ ВЕТВЬ R-M222.
Удивительной, только не по широте ареала, а как раз по его компактности, можно считать гаплогруппу R-M222 (рис. 2.17). Распространение этой гаплогруппы практически исключительно на Британских островах наводит на предположение о ее местном происхождении. Согласно оценкам www.yfull.com, экспансия гаплогруппы R-M222 датируется всего лишь 1,8 тыс. лет назад (в интервале 1,3-2,4 тыс. лет назад). Иными словами, гаплогруппа появилась в популяциях Британских островов во 2 тыс. до н.э., а миграции, которые распространили её, происходили в начале нашей эры, то есть примерно в римское время.
Пример филогеографии гаплогруппы R-L265 показывает, сколь подробная генетическая информация скрывается в пределах каждой гаплогруппы Y-хромосомы. Остается надеяться, что по мере накопления данных по полным геномам будут обнаружены информативные маркеры и в пределах других гаплогрупп.
ИНДИЙСКАЯ ГАПЛОГРУППА R2 (R-L266).
Гаплогруппа R2 (R-L266) происходит из того же корня, что и гаплогруппа R1, давшая начало гаплогруппам R1a и R1b. Карта распространения гаплогруппы R2 (R-L266) (рис. 2.18) показывает, что для народонаселения Европы в целом R2 не характерна: она встречается с крайне низкими частотами (менее 3%) лишь на севере Аппенинского полуострова и в Сардинии, у популяции турок вблизи пролива Дарданеллы, в центральной Анатолии и Закавказье. С чуть более высокой частотой 5% гаплогруппа R2 (R-L266) замечена в юго-восточной Турции, с частотой 6% — у калмыков. Такая скудная география гаплогруппы в пределах картографируемого ареала, пожалуй, неудивительна: ведь основной ареал R2 (R-L266) охватывает далекие территории Индийского полуострова и Центральной Азии, а с низкими частотами гаплогруппа доходит до Китая на востоке и до Юго-Западной Азии — на западе своего ареала.
 Y-хромосомы в Европе.") Рис. 2.18. География гаплогруппы R-L266 (R2) Y-хромосомы в Европе.
ГАПЛОГРУППА I И ЕЕ СУБВАРИАНТЫ
Продолжая тему о высокой информативности выделения субтипов и анализа их географического распределения, напомню ставшую классической работу [Rootsi et al., 2004], в которой гаплогруппа I-М170 была филогенетически подразделена на три субтипа. Оказалось, что эти субтипы имеют совершенно различную географическую приуроченность: гаплогруппа I1-М253 (на картах — I-L118) приурочена к северной Европе, I2a-Р37 – к Балканскому региону, а I2b-М223 (на картах - I-L35) локализована преимущественно в Северо-Западной Европе. Именно после этой работы и ряда ей подобных стало ясно, что обнаружение новых SNP маркеров, подразделяющих гаплогруппу на субветви, является магистральным путем повышения информативности анализа Y-хромосомы. Но долгое время движение по этому пути было медленным, и лишь после внедрения полного секвенирования Y-хромосомы новые маркеры стали накапливаться лавинообразно. Так, в течение долгого времени не удавалось обнаружить новые SNP маркеры в пределах гаплогруппы I, или же новые открываемые SNP маркеры не выделяли новых ветвей с четкой географией. Например, в работе [Underhill et al., 2008] были обнаружены новые маркеры и реорганизована топология ветвей гаплогруппы I-М170, но сохранились, пусть под новыми названиями, те же три основные ветви, а новые обнаруженные варианты были редкими и не характеризовались четкими географическими трендами.
«СКАНДИНАВСКАЯ» ГАПЛОГРУППА I-L118.
Карта распространения «скандинавской» гаплогруппы I-L118 (Рис. 2.19) показывает ее максимальные частоты не только в географической Скандинавии, но и на территориях, тесно связанных с ней исторически: Дании, Шотландии, западных районах Финляндии. Поэтому любопытна область умеренных частот у русских популяций к северу от Волги, выделяющаяся на фоне низких частот у большинства славянских популяций. Нельзя исключать, что это отражает исторические контакты с норманнскими популяциями. Хотя эти контакты были более интенсивны для северо-западных русских популяций (на пути «из варяг в греки»), но территории северо-запада были сравнительно густо населены, поэтому влияние «варягов» могло оказаться более значимым для меньших по размеру популяций Заволжья.
 Рис. 2.19. География гаплогруппы I-L118 Y-хромосомы в Европе.
«БАЛКАНСКАЯ» ГАПЛОГРУППА I-Р37
Карта распространения «балканской» гаплогруппы I-Р37 (Рис. 2.20) показывает ее высокие частоты также и в Италии. В целом, эта гаплогруппа распространена практически по всей Европе, кроме Скандинавии. В Восточной Европе плавный градиент убывания ее частоты с юго-запада (с Украины) к северу и востоку очень напоминает карту первой главной компоненты по классическим генетическим маркерам [Рычков и др., 2002]. Также стоит отметить достаточно выраженное совпадение ареала этой гаплогруппы с географическими границами Европы – при высокой частоте в Причерноморье гаплогруппа очень редка на Северном Кавказе и почти отсутствует в Закавказье, а при максимальных частотах на Балканах очень редка в соседней Малой Азии.
 Рис. 2.20. География гаплогруппы I-P37 Y-хромосомы в Европе.
«ВАРЯГО-ГРЕЧЕСКАЯ» ГАПЛОГРУППА I-L35
Вдоль берегов Северного и Балтийского морей, и немного вглубь от них — так можно было бы описать географию гаплогруппы I-L35, если бы еще не два дополнения на северо-востоке и юго-востоке. Хотя максимум частоты гаплогруппы I-L35 (17%) отмечен в Швеции (рис. 2.21), зона более редкой встречаемости гаплогруппы (3-7%) охватывает северную часть Центральной Европы, на юго-востоке отдельной областью очень низких частот (1-3%) охватывает Балканский полуостров и западную часть Малой Азии, а на северо-востоке проявляется у русских популяций Тверской и Костромской областей. Можно заметить, что ареал гаплогруппы I-L35 — довольно умеренный и однородный по значениям частот гаплогруппы (на большинстве ареала частота варьирует от 2-3% до 7%, только пик в Швеции до 17%) — связывает несколько крайних областей Европы: Скандинавию, северные районы Восточной Европы и восток Южной Европы. Вполне возможно, что география гаплогруппы I-L35 действительно отражает один из генетических следов торговых путей между севером и югом Европы. Не менее вероятно, что каждая из этих зон связана со своим субвариантом этой гаплогруппы.
 Рис. 2.21. География гаплогруппы I-L35 Y-хромосомы в Европе.
ГАПЛОГРУППА N И ЕЕ СУБВАРИАНТЫ
Гаплогруппа N в пределах Европейского региона представлена двумя большими ветвями — гаплогруппами N1c и N1b (далее и на картах N-M46 и N-P43 соответственно). Хотя общие ареалы этих двух гаплогрупп в Европе пересекаются, зоны их максимальных частот разграничиваются на севере рекой Печорой, а южнее — Уральским хребтом: с запада этой условной «границы» остаются зоны повышенной встречаемости гаплогруппы N-M46, а с востока — зона максимальных частот N-P43. Рассмотрим географию этих двух «братских» ветвей гаплогруппы N чуть подробнее.
СЕВЕРОЕВРОПЕЙСКАЯ ЗАПАДНАЯ ГАПЛОГРУППА N-M46
Карта распространения гаплогруппы N-М46 (Рис. 2.22) показывает ее максимальные частоты у финнов. Ее частоты высоки и у других популяций северо-востока Европы. Хотя в целом эта гаплогруппа оказывается приуроченной к финно-угорским популяциям, но она столь же часта и у северных русских (что, по-видимому, объясняется ассимиляцией дославянского населения, как будет описано в следующих главах). Наиболее любопытным представляется область высокой частоты этой гаплогруппы у балтов (латышей и литовцев). В данном случае генетические границы хорошо совпадают с лингвистическими: граница ареалов балтов и западных славян совпадает с резко выраженным «обрывом» частоты гаплогруппы. Похожая картина наблюдается и на границе с белорусами.
 Y-хромосомы в Европе.") Рис. 2.22. География гаплогруппы N-M46 (N1c) Y-хромосомы в Европе.
«СЕВЕРОУРАЛЬСКАЯ» ГАПЛОГРУППА N-P43
Карта распространения гаплогруппы N-P43 (Рис. 2.23) выявляет также северный, но куда более узкий ареал. Максимальные частоты этой гаплогруппы наблюдаются на крайнем северо-востоке Европы и плавно переходят в мировой максимум этой гаплогруппы в Западной Сибири. С умеренными частотами эта гаплогруппа распространена по всему Уралу и Приуралью. Заметен плавный градиент убывании частоты к западу и югу, так что у популяций Центральной России и нижней Волги эта гаплогруппа еще встречается (хотя и с минимальным частотами), а у других популяций Европы практически полностью отсутствует.
 Y-хромосомы в Европе.") Рис. 2.23. География гаплогруппы N-P43 (N1b) Y-хромосомы в Европе.
ГАПЛОГРУППА E И ЕЕ СУБВАРИАНТЫ
Большинство вышеописанных гаплогрупп в основном характеризовали условно северную половину Европейского континента. Рассматривая географию следующей гаплогруппы — E – мы переместим внимание на юг, к берегам Средиземного моря. В Европе встречаются четыре основные ветви гаплогруппы E: E-L142 и E-V22 (восходящие к общему корню — гаплогруппе E-M78), E-M81 и E-M123.
ВОСТОЧНО-СРЕДИЗЕМНОМОРСКАЯ ГАПЛОГРУППА E-L142
Карта гаплогруппы E-L142 (Рис. 2.24) отчасти напоминает карту гаплогруппы I-Р37 (рис. 2.20) – максимальные частоты обеих этих гаплогрупп приурочены к Балканскому полуострову. Также очень похожи их паттерны: убывание частоты к северу, западу и востоку. Основные различия между трендами двух этих гаплогрупп состоят, во-первых, в меньшей частоте E-L142 (на большинстве территорий она «уступает» I-Р37 на один интервал шкалы карты) и, во-вторых, в распространении E-L142 не только в Европе, но и по всему Средиземноморью, включая Ближний Восток и северные окраины Африки. В пределах Балкан I-Р37 особенно часта у боснийцев и хорватов, а E-L142 преобладает южнее – у сербов, албанцев и греков.
 Рис. 2.24. География гаплогруппы E-L142 Y-хромосомы в Европе.
«ЕГИПЕТСКАЯ» ГАПЛОГРУППА E-V22
Два небольших «глазка» на севере и юге Пиренейского полуострова и Аппенинский полуостров – территории с частотой гаплогруппы E-V22 до 5%, небольшая область на северо-западе Малой Азии (в окрестностях Стамбула) с частотой до 6%: на этом можно было бы закончить описание географии гаплогруппы E-V22 в географических границах Европы (рис. 2.25). Однако география гаплогруппы E-V22 в Европе в основном следует Средиземноморскому побережью, поэтому стоит проследить её распространение и южнее. Карта на рис. 2.25 показывает плавное повышение частоты гаплогруппы E-V22 в двух противолежащих регионах: на юго-западе и юго-востоке Средиземноморья. В юго-западном Средиземноморье у берегов Гибралтарского пролива и вглубь Марокко частота гаплогруппы E-V22 вырастает до 7%. В юго-восточном Средиземноморье повышение частоты гаплогруппы E-V22 отмечается от Малой Азии на юг — через Левант вплоть до дельты Нила, где она достигает 14%, и ещё южнее вдоль Нила, где у египетских арабов оазиса Бахария она достигает мирового максимума 22%. Из этих двух неевропейских регионов распространения гаплогруппы E-V22, вероятно, именно последний (с более высокими частотами и широким распространением) послужил основным источником распространения гаплогруппы E-V22 в Европу.
 Рис. 2.25. География гаплогруппы E-V22 Y-хромосомы в Европе.
САХАРСКАЯ ГАПЛОГРУППА E-M81
Если две предыдущих ветви гаплогруппы E – E-L142 и E-V22 – демонстрировали в основном распространение вдоль северных и восточных берегов Средиземного моря, иногда с углублением на север и восток континента, то география третьей ветви — гаплогруппы E-M81- больше следует южному Средиземноморью. Карта распространения гаплогруппы E-M81 на рис. 2.26 хотя и показывает заметное присутствие гаплогруппы на Ближнем Востоке (частота до 20%), однако основная зона высоких частот находится в северо-западной Африке. Из этого максимума отмечено плавное снижение частоты в северном направлении через Гибралтарский пролив с умеренным распространением (2-10%) гаплогруппы E-M81 на территории Пиренейского полуострова и юго-западной Франции, узкие области низких частот (не более 3%) на Аппенинском и Балканском полуостровах, а также северо-востоке Малой Азии.
 Рис. 2.26. География гаплогруппы E-M81 Y-хромосомы в Европе.
 Рис. 2.27. География гаплогруппы E-M123 Y-хромосомы в Европе.
ПЕРЕДНЕАЗИАТСКАЯ ГАПЛОГРУППА E-M123
Гаплогруппа E-M123 достигает максимума в Передней Азии (20-25%), ближе к Европе (в Малой Азии) снижается до 15%. В самой же Европе она с низкими частотами распространена вдоль северных берегов Средиземного моря, и отдельными вкраплениями проявляется в Центральной Европе (рис. 2.27).
ГАПЛОГРУППА G И ЕЕ СУБВАРИАНТЫ
Карты распространения гаплогруппы двух ветвей гаплогруппы G-P15 – гаплогрупп G-P303 и G-P16 – в целом имеют похожее распространение и даже в общеевропейском масштабе карт видно, что обе приурочены в первую очередь к Кавказу. Однако своих максимальных частот они достигают в разных частях Кавказа региона: гаплогруппа G-P303 преобладает на крайнем северо-западе у причерноморских шапсугов, а гаплогруппа G-P16 — на Центральном Кавказе у осетин-иронцев (рис. 2.28 и рис. 2.29). Это полностью подтверждается при отдельном изучении генофонда Кавказа [Balanovsky et al., 2011; глава 7 этой книги].
 Рис. 2.28. География гаплогруппы G-P303 Y-хромосомы в Европе.
ЗАПАДНОКАВКАЗСКАЯ ГАПЛОГРУППА G-P303
Карта показывает, что гаплогруппа G-P303 (Рис. 2.28) также распространена на Ближнем Востоке, а в Европе встречается в основном в ее юго-восточных областях. Зоны высокой частоты гаплогруппы G-P303 тянутся вдоль восточного побережья Черного моря и приурочены к популяциям народов Западного Кавказа, в основном, абхазо-адыгской лингвистической группы. С меньшими частотами (до 10%) гаплогруппа G-P303 распространена в Северном Причерноморье (на территории Крымского полуострова и в Приазовье), область низких частот (3-5%) тянется далее на север в зону украинской лесостепи. Локальное повышение частоты гаплогруппы G-P303 до 13% также наблюдается в Поволжье у популяции мордвы мокша и окружается областью низких частот.
 Рис. 2.29. География гаплогруппы G-P16 Y-хромосомы в Европе.
ЦЕНТРАЛЬНОКАВКАЗСКАЯ ГАПЛОГРУППА G-P16
География гаплогруппы G-P16 (рис. 2.29) более узкая по сравнению с её «братской» ветвью G-P303. Она охватывает исключительно Кавказский регион и Малую Азию. В пределах этого ареала частота гаплогруппы G-P16 резко падает от максимальных значений у осетин (73% у осетин-иронцев и 56% у осетин-дигорцев) к значениям в диапазоне 10-20% у народов Северо-Западного Кавказа (абхазов, черкесов, балкарцев и карачаевцев) и затем снижается до минимальной отметки у турок, армян и азербайджанцев.
ГАПЛОГРУППА J И ЕЕ СУБВАРИАНТЫ
Гаплогруппа J представлена двумя крупными ветвями: J-L255 (J1) и J-L228 (J2), из которых в Европе большее распространение получила вторая. Хотя обе эти ветви по своему происхождению ближневосточные и отражают древние взаимосвязи населения Европы и жителей намного более южных регионов Евразии, имеет смысл рассмотреть отдельно географию и каждой из этих линий в целом, и каждой из внутренних ветвей более распространенной в Европе гаплогруппы J-L228 (J2).
 Y-хромосомы в Европе.") Рис. 2.30. География гаплогруппы J-L255 (J1) Y-хромосомы в Европе.
ГАПЛОГРУППА J-L255 (J1): от Передней Азии к пикам Восточного Кавказа
Распространение гаплогруппы J-L255 (Рис. 2.30) в картографируемом ареале охватывает преимущественно его юго-восточные области: Кавказ, Малую Азию, Северное Причерноморье, Балканский полуостров; небольшими зонами – юг Аппенинского полуострова и Франции, юго-запад Пиренейского полуострова. В пределах региона максимальных частот – Кавказа – гаплогруппа J-L255 выделяет народы Восточного Кавказа, где на ёё долю приходится львиная доля всего генофонда. Максимума гаплогруппа J-L255 достигает в Дагестане (у кубачинцев, даргинцев, табасаранцев и аварцев ее частоты колеблются от 63 до 98%), и снижается до 44% у лезгин и 21% у чеченцев. На территории Малой Азии доля гаплогруппы J-L255 составляет от 5% до 15%.
ГАПЛОГРУППА J-L228 (J2) В ЦЕЛОМ
Карта распространения гаплогруппы J-L228 (J2) рисует иную картину (рис. 2.31). В отличие от большинства рассмотренных выше гаплогрупп, J-L228 (J2) распространена с высокими частотами не в Европе, а на Ближнем Востоке и в Северной Африке. Конечно, ее ареал заходит и в соседние с этими регионами южные области Европы: Испанию (особенно в ее южные районы), Италию (также особенно южную), юг Балканского полуострова. Но максимальных частот (красный цвет на карте) эта гаплогруппа достигает в некоторых популяциях Северного Кавказа. Как было показано в нашем исследовании [Balanovsky et al., 2011; глава 7 данной книги], а также в работе [Yunusbaev et al., 2012] кавказский максимум приурочен к популяциям нахской группы (чеченцам и ингушам).
 Y-хромосомы в Европе.") Рис. 2.31. География гаплогруппы J-L228 (J2) Y-хромосомы в Европе.
 Рис. 2.32. География гаплогруппы J-M67 Y-хромосомы в Европе
ГАПЛОГРУППА J-L152 И ЕЁ СУБВАРИАНТ J-M67
География гаплогруппы J-L152 на территории Европы довольно обширна: зона низких частот тянется с северо-запада Балканского полуострова через Центральную Европу и до Пиренейского полуострова. Зона повышенных частот (от 15%) охватывает Малую Азию и Кавказ. Практически повторяет эту закономерность, но несколько в более узком географическом охвате и в целом с более низкими частотами, распространение ее «сыновней» ветви – гаплогруппы J-M67 (рис. 2.32). Однако при этом гаплогруппа J-M67 по своей частоте больше приурочена к Кавказу, составляя на территории Малой Азии не более 13% генофонда.
ГАПЛОГРУППА J-L282
Распространение гаплогруппы J-L282 в Европе (рис. 2.33) заметно отличается от предыдущей ветви. На фоне обширной зоны низких частот выделяются три области, где доля гаплогруппы J-L282 в генофонде повышена до 10-15%: север Фенноскандии (правда, этот максимум опирается лишь на данные по одной популяции), Поволжье (популяция мордвы мокша) и север Балканского полуострова (несколько популяций албанцев).
 Рис. 2.33. География гаплогруппы J-L282 Y-хромосомы в Европе.
РЕДКИЕ ГАПЛОГРУППЫ L, Q, T
ГАПЛОГРУППА L-M11
Гаплогруппа L-M11 (рис. 2.34) в Европе приурочена к юго-восточным регионам (за исключением единичного случая в Бельгии): Крымский полуостров, европейская часть Турции и Северный Кавказ. При этом частота гаплогруппы L-M11 плавно увеличивается в юго-восточном направлении, достигая 14% на территории Восточного Кавказа у чеченцев и 13% на северо-востоке Малой Азии у турок. Далее ее частота повышается на Ближнем и Среднем Востоке, достигая индийского максимума уже далеко за пределами картографируемого ареала.
 Рис. 2.34. География гаплогруппы L-M11 Y-хромосомы в Европе.
ГАПЛОГРУППА Q-M242
Гаплогруппа Q-M242 в Европе практически не встречается, за исключением нескольких точек с низкими частотами (до 3%) в Центральной и Восточной Европе, на Крымском полуострове и в восточной половине Малой Азии (рис. 2.35). Плавным коридором от северо-западного побережья Каспийского моря далее на восток с постепенным ростом частоты ареал гаплогруппы Q-M242 переходит в Центральную Азию и Сибирь, где она достигает своего максимума.
 Рис. 2.35. География гаплогруппы Q-M242 Y-хромосомы в Европе.
 Рис. 2.36. География гаплогруппы T-L206 Y-хромосомы в Европе.
ГАПЛОГРУППА T
Частота гаплогруппы T-L206 в народонаселении Европы не превышает 5% (рис. 2.36). Ее ареал в Европе лоскутный и сводится на западе к трем узким областям на Пиренейском полуострове и в Сардинии, а на востоке – к зоне в Северном Причерноморье с переходом на северо-восток в украинскую лесостепь, единичными случаями в Поволжье у казанских татар и на Северном Кавказе у кубанских ногайцев. За пределами Европы частота гаплогруппы Т столь же невелика, но зато ее ареал становится уже не лоскутным, а почти непрерывным, охватывая всю Малую Азию, Ближний Восток и северо-восток Африки.
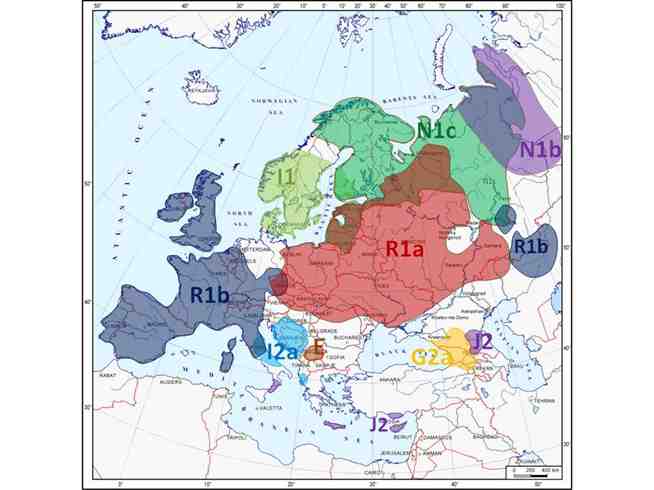
Y-ГЕНОФОНД ЕВРОПЫ — ПРИНЦИП ПАЗЗЛОВ
Таким образом, проведенное исследование изменчивости Y-хромосомы в популяциях Европы, основанное на объединении обширных собственных данных с литературными, подтвердило и уточнило вывод о том, что основной чертой структурированности генофонда является четкое выделение географических зон, в каждой из которых преобладает своя гаплогруппа. Это убедительно демонстрируют карты распределения гаплогрупп (рис. 2.2.-2.36).
Однако при размещении данных по каждой гаплогруппе на отдельной карте трудно отследить перекрывание – или же, напротив, не перекрывание — ареалов гаплогрупп. Поэтому мы совместили зоны распространения всех гаплогрупп на одной карте (рис. 2.37). На этом рисунке контур каждой из девяти основных европейских гаплогрупп строго соответствует территории, на которой частота этой гаплогруппы выше 35%, т.е. более трети генофонда. Можно видеть, что каждая гаплогруппа действительно занимает свой собственный ареал. Области, остающиеся белыми, невелики – это те территории, на которых разнообразие гаплогрупп высоко и ни одна гаплогруппа не достигает уровня 35%.
Полученная карта ярко демонстрирует «принцип паззлов» — высокую географическую специфичность гаплогрупп Y-хромосомы. Благодаря этому на каждой территории, хотя и присутствует много гаплогрупп, но доминируют лишь одна-две. Таким образом генофонд Европы оказывается составлен, как из паззлов, из ареалов преимущественного распространения разных гаплогрупп.
 Рис. 2.37. Структура генофонда Европы по Y-хромосоме: «принцип паззлов».
В каждой географической части Европы доминирует одна гаплогруппа, редко встречающаяся в остальных частях. Карта показывает, к каким частям Европы приурочена каждая из основных гаплогрупп. Карта основана на точных значениях частот. Цветом показаны зоны с частотой гаплогруппы выше порогового значения 0,35 (то есть на выделенных цветом территориях более трети генофонда относится к данной гаплогруппе).
[1] Отметим в скобках, что объективная приуроченность данной гаплогруппы к славянским популяциям на данной весьма ограниченной территории, к сожалению, стала одним из поводов к столь распространенному на любительских форумах в интернете навешиванию «славянского» ярлыка на всю гаплогруппу R1a. Но такая привязка гаплогруппы — на всем протяжении ее евразийского ареала в пространстве и несоизмеримого со славянством протяженного существования во времени — к одной лишь славянской лингвистической группе, увы, уже не только граничит со лженаукой, но и переходит эту грань.
Похожие статьи
| Публикуем фрагмент из книги О.П. Балановского "Генофонд Европы" (выйдет в декабре 2015 г.). Карты генетических расстояний позволят своими глазами увидеть, насколько генофонд отдельного народа похож на все остальные генофонды Европы. Представлены карты первой из шести серий - "Народы Северо-Восточной Европы": от карел и вепсов, от эстонцев и коми, от литовцев и латышей, от северных русских и финнов. | Следующий фрагмент из книги О.П. Балановского «Генофонд Европы» описывает своеобразие генофондов западных и восточных славян. Карты генетических расстояний обобщают разнообразие гаплогрупп Y-хромосомы и позволяют самим убедиться, насколько каждая точка в ареале Европы генетически близка к средним параметрам каждого из народов западных и восточных славян: их генофонды оказались настолько близки, что им хочется дать имя "генофонд северных славян". | Эта серия карт очередного фрагмента из книги О.П. Балановского «Генофонд Европы» описывает разнообразие Y-хромосомного генофонда Волжско-Уральского региона. Рассмотрена только полоса соседствующих популяций - Башкортостана, Татарстана, Чувашии и Мордовии. Но несмотря на их относительно небольшой суммарный ареал, генофонды оказались своеобразны и даже загадочны.
| Публикуем четвертую серию карт генетических расстояний на основе гаплогрупп Y-хромосомы из книги О.П. Балановского «Генофонд Европы». Эти карты отражают генетический ландшафт северной окраины Балкан, где проживают разноязыкие народы, говорящие на языках трех лингвистических семей. | | В пятой серии карт (из книги О. Балановского «Генофонд Европы») мы видим степень близости к каждой из популяций Европы южных славян - македонцев, сербов, хорватов, боснийцев и герцеговинцев. Географически их объединяет принадлежность к Балканам, а генетическое своеобразие связывается с сохранением субстратного генофонда тех балканских племен и народов, которые стали говорить на славянских языках. | Последняя серия карт генетических расстояний (из книги О. Балановского «Генофонд Европы») от народов, ничем друг на друга не похожих – ни языком, ни географией. Но зато эти три генофонда окаймляют пространство народов, рассмотренных в пяти предыдущих сериях, и позволяют увидеть, насколько велики различия генофондов европейской окраины Евразии. Эти три этноса – албанцы, шведы, ногайцы - не только географически «расставлены» по трем «концам земли», но и генетически полярно различны, показывая масштаб разнообразия генофонда Европы. |
|
Избранное
Анализ древних геномов с запада Иберийского полуострова показал увеличение генетического вклада охотников-собирателей в позднем неолите и бронзовом веке. След степной миграции здесь также имеется, хотя в меньшей степени, чем в Северной и Центральной Европе.
|
Геологи показали, что древний канал, претендующий на приток мифической реки Сарасвати, пересох еще до возникновения Индской (Хараппской) цивилизации. Это ставит под сомнение ее зависимость от крупных гималайских рек.
|
Текст по пресс-релизу Института археологии РАН о находке наскального рисунка двугорбого верблюда в Каповой пещере опубликован на сайте "Полит.ру".
|
На основе изученных геномов бактерии Yersinia pestis из образцов позднего неолита – раннего железного века палеогенетики реконструировали пути распространения чумы. Ключевое значение в ее переносе в Европу они придают массовой миграции из причерноморско-каспийских степей около 5000 лет назад. По их гипотезе возбудитель чумы продвигался по тому же степному коридору с двусторонним движением между Европой и Азией, что и мигрирующее население.
|
Генетическое разнообразие населения Сванетии в этой работе изучили по образцам мтДНК и Y-хромосомы 184 человек. Данные показали разнообразие митохондриального и сравнительную гомогенность Y-хромосомного генофонда сванов. Авторы делают вывод о влиянии на Y-хромосомный генофонд Южного Кавказа географии, но не языков. И о том, что современное население, в частности, сваны, являются потомками ранних обитателей этого региона, времен верхнего палеолита.
|
Опубликовано на сайте Коммерсант.ru
|
Авторы свежей статьи в Nature опровергают представления о почти полном замещении охотников-собирателей земледельцами в ходе неолитизации Европы. Он и обнаружили, что генетический вклад охотников-собирателей различается у европейских неолитических земледельцев разных регионов и увеличивается со временем. Это говорит, скорее, о мирном сосуществовании тех и других и о постоянном генетическом смешении.
|
Последние дни у нас веселые – телефон звонит, не переставая, приглашая всюду сказать слово генетика. Обычно я отказываюсь. А здесь все одно к одному - как раз накануне сдали отчет на шестистах страницах, а новый – еще только через месяц. И вопросы не обычные - не про то, когда исчезнет последняя блондинка или не возьмусь ли я изучить геном Гитлера. Вопросы про президента и про биологические образцы.
|
В Медико-генетическом научном центре (ФГБНУ МГНЦ) 10 ноября прошла пресс-конференция, на которой руководители нескольких направлений рассказали о своей работе, связанной с генетическими и прочими исследованиями биологических материалов.
|
Горячая тема образцов биоматериалов обсуждается в программе "В центре внимания" на Радио Маяк. В студии специалисты по геногеографии и медицинской генетике: зав. лаб. геномной географии Института общей генетики РАН, проф. РАН Олег Балановский и зав. лаб. молекулярной генетики наследственных заболеваний Института молекулярной генетики РАН, д.б.н., проф. Петр Сломинский.
|
О совсем недавно открытой лейлатепинской культуре в Закавказье, ее отличительных признаков и корнях и ее отношениях с известной майкопской культурой.
|
Интервью О.П.Балановского газете "Троицкий вариант"
|
В издательстве «Захаров» вышла книга «Эта короткая жизнь: Николай Вавилов и его время». Ее автор Семен Ефимович Резник, он же автор самой первой биографической книги о Н.И.Вавилове, вышедшей в 1968 году в серии ЖЗЛ.
|
Исследование генофонда четырех современных русских популяций в ареале бывшей земли Новгородской позволяет лучше понять его положение в генетическом пространстве окружающих популяций. Он оказался в буферной зоне между северным и южным «полюсами» русского генофонда. Значительную (пятую) часть генофонда население Новгородчины унаследовало от финноязычного населения, которое, видимо, в свою очередь, впитало мезолитический генофонд Северо-Восточной Европы. Генетические различия между отдельными популяциями Новгородчины могут отражать особенности расселения древних славян вдоль речной системы, сохранившиеся в современном генофонде вопреки бурным демографическим событиям более поздних времен.
|
На "Эхе Москвы" в программе "Культурный шок" беседа глав. ред. Алексея Венедиктова с д.б.н., зав. кафедрой биологической эволюции Биологического факультета МГУ Александром Марковым.
|
О том, неужели кто-то пытается придумать биологическое оружие против граждан России — материал Марии Борзуновой (телеканал "Дождь").
|
Отличная статья на сайте "Московского комсомольца"
|
Что такое биоматериал? Где он хранится и как используется? Об этом в эфире “Вестей FM” расскажут директор Института стволовых клеток человека Артур Исаев и заведующий лабораторией геномной географии Института общей генетики имени Вавилова, доктор биологических наук, профессор РАН Олег Балановский.
|
Что стоит за высказыванием В.В.Путина о сборе биологических материалов россиян, и реакцию на его слова в студии "Радио Свобода" обсуждают: политик Владимир Семаго, доктор биологических наук, генетик Светлана Боринская, руководитель лаборатории геномной географии Института общей генетики РАН Олег Балановский.
|
Как сказал ведущий программы «Блог-аут» Майкл Наки, одна из самых обсуждаемых новостей недели – это высказывание Владимира Путина, про то, что собираются биоматериалы россиян – массово и по разным этносам. И это было бы смешно, когда бы не было так грустно - если бы после этого высказывания всякие каналы не начали выпускать сюжеты о биооружии, которое готовится против россиян. По поводу этой странной истории ведущий беседует с д.б.н., проф. РАН О.П.Балановским.
|
Ведущие специалисты в области генетики человека считают напрасными страхи перед неким «этническим оружием». Сделать его невозможно.
|
Комментируем ситуацию вокруг вопроса Президента РФ, кто и зачем собирает биологический материал россиян.
|
В африканских популяциях, как выяснилось, представлено большое разнообразие генетических вариантов, отвечающих за цвет кожи: не только аллели темной кожи, но и аллели светлой кожи. Последних оказалось особенно много у южноафриканских бушменов. Генетики пришли к заключению, что варианты, обеспечивающие светлую кожу, более древние, и возникли они в Африке задолго до формирования современного человека как вида.
|
Анализ генома 40-тысячелетнего человека из китайской пещеры Тяньянь показал его генетическую близость к предкам восточноазиатских и юговосточных азиатских популяций и указал на картину популяционного разнообразия в верхнем палеолите. Исследователи полагают, что 40-35 тыс. лет назад на территории Евразии обитали не менее четырех популяций, которые в разной степени оставили генетический след в современном населении.
|
В Санкт-Петербургском государственном университете, в Петровском зале здания Двенадцати коллегий состоялись чтения, посвященные 90-летию со дня рождения Льва Самуиловича Клейна. Большинство из выступавших на них археологов, антропологов, историков и других специалистов считают себя его учениками, которым он привил основы научного мышления, научил идти непроторенными дорогами, показал пример преодоления обстоятельств и стойкости в борьбе. Научные доклады начинались со слов признательности учителю. Представляем здесь выступление доктора исторических наук, профессора СПбГУ, главного научного сотрудника Музея антропологии и этнографии РАН Александра Григорьевича Козинцева.
|
Накануне 110-летия со дня рождения знаменитого антрополога и скульптора, автора всемирно известного метода реконструкции лица по черепу Михаила Михайловича Герасимова, в Дарвиновском музее прошел вечер его памяти. О том, как появился знаменитый метод, о работах мастера и развитии этого направления в наши дни рассказали его последователи и коллеги.
|
Генетики секвенировали митохондриальную ДНК 340 человек из 17 популяций Европы и Ближнего Востока и сравнили эти данные с данными по секвенированию Y-хромосомы. Демографическая история популяций, реконструированная по отцовским и материнским линиям наследования, оказалась совершенно разной. Если первые указывают на экспансию в период бронзового века, то вторые хранят память о расселении в палеолите после окончания оледенения.
|
Анализ геномов четырех индивидов с верхнепалеолитической стоянки Сунгирь показал, что они не являются близкими родственниками. Из этого авторы работы делают вывод, что охотники-собиратели верхнего палеолита успешно избегали инбридинга, так как каждая группа была включена в разветвленную сеть по обмену брачными партнерами.
|
Изучив 16 древних геномов из Африки возрастом от 8100 до 400 лет, палеогенетики предлагают картину смешений и перемещений, приведшую к формированию современных африканских популяций.
|
Анализ семи древних геномов из Южной Африки показал глубокие генетические различия между бушменами и прочими африканскими и неафриканскими популяциями. Время формирования первой развилки на древе человечества соответствует периоду формирования современного человека как вида, авторы оценили его в диапазоне от 350 до 260 тысяч лет назад.
|
Генетический ландшафт Папуа Новая Гвинея отмечен кардинальными различиями между горными и равнинными популяциями. Первые, в отличие от вторых, не обнаруживают влияния Юго-Восточной Азии. Среди горных популяций отмечается высокое генетическое разнообразие, возникшее в период возникновения земледелия. Делается вывод, что неолитический переход не всегда приводит к генетической однородности населения (как в Западной Евразии).
|
В неолитизации Европы роль культурной диффузии была очень незначительной. Основную роль играло распространение земледельцев с Ближнего Востока, которые почти полностью замещали местные племена охотников-собирателей. Доля генетического смешения оценивается в 2%. К таким выводам исследователей привел анализ частоты гаплогрупп митохондриальной ДНК и математическое моделирование.
|
Сочетание генетического и изотопного анализа останков из захоронений на юге Германии продемонстрировало патрилокальность общества в позднем неолите – раннем бронзовом веке. Мужчины в этом регионе вели оседлый образ жизни, а женщины перемещались из других регионов.
|
Наш постоянный читатель и активный участник дискуссий на сайте Лев Агни поделился своим мнением о том, что противопоставить изобилию некачественных научных публикаций в области истории.
|
Древние геномы изучили по аллелям, ассоциированным с болезнями, и вычислили генетический риск наших предков для разных групп заболеваний. Оказалось, что этот риск выше у более древних индивидов (9500 лет и старше), чем у более молодых (3500 лет и моложе). Обнаружилась также зависимость генетического риска заболеваний от типа хозяйства и питания древних людей: скотоводы оказались более генетически здоровыми, чем охотники-собиратели и земледельцы. Географическое местоположение лишь незначительно повлияло на риск некоторых болезней.
|
Международная группа археологов опровергла датировку выплавки меди в Чатал-Хююке – одном из самых известных поселений позднего неолита в центральной Турции. Статья с результатами исследования опубликована в журнале Journal of Archaeological Science .
|
В продолжение темы майкопской культуры перепечатываем еще одну статью археолога, канд. ист. наук Н.А.Николаевой, опубликованную в журнале Вестник Московского государственного областного университета (№1, 2009, с.162-173)
|
В продолжение темы, рассмотренной в статье А.А.Касьяна с лингвистических позиций, и с разрешения автора перепечатываем статью археолога, к.и.н. Надежды Алексеевны Николаевой, доцента Московского государственного областного университета. Статья была опубликована в 2013 г. в журнале Восток (Оriens) № 2, С.107-113
|
Частичный перевод из работы Алексея Касьяна «Хаттский как сино-кавказский язык» (Alexei Kassian. 2009–2010. Hattic as a Sino-Caucasian language. Ugarit-Forschungen 41: 309–447)
|
Несмотря на признание исследований по географии генофондов со стороны мирового научного сообщества и все возрастающую роль геногеографии в междисциплинарных исследованиях народонаселения, до сих пор нет консенсуса о соотношении предметных областей геногеографии и этнологии. Генетики и этнологи часто работали параллельно, а с конца 2000-х годов началось их тесное сотрудничество на всех этапах исследования – от совместных экспедиций до совместного анализа и синтеза. Приведены примеры таких совместных исследований. Эти примеры демонстрируют, что корректно осуществляемый союз генетики и этнологии имеет добротные научные перспективы.
|
Генетический анализ показал, что население Мадагаскара сформировалось при смешении предков африканского происхождения (банту) и восточноазиатского (индонезийцы с Борнео). Доля генетических компонентов разного происхождения зависит от географического региона: африканского больше на севере, восточноазиатского – на юго-востоке. На основании картины генетического ландшафта авторы реконструируют историю заселения Мадагаскара – переселенцы из Индонезии появились здесь раньше, чем африканцы.
|
Появились доказательства того, что анатомически современный человек обитал на островах Индонезии уже в период от 73 до 63 тыс. лет назад, статья с результатами этой работы опубликована в Nature.
|
Анализ геномов бронзового века с территории Ливана показал, что древние ханаанеи смешали в своих генах компоненты неолитических популяций Леванта и халколитических - Ирана. Современные ливанцы получили генетическое наследие от ханаанеев, к которому добавился вклад степных популяций.
|
В журнале European Journal of Archaeology опубликована дискуссия между проф. Л.С.Клейном и авторами статей в Nature (Haak et al. 2015; Allentoft 2015) о гипотезе массовой миграции ямной культуры по данным генетики и ее связи с происхождением индоевропейских языков. Дискуссия составлена из переписки Л.С.Клейна с несколькими соавторами (Вольфганг Хаак, Иосиф Лазаридис, Ник Пэттерсон, Дэвид Райх, Кристиан Кристиансен, Карл-Гёран Шорген, Мортен Аллентофт, Мартин Сикора и Эске Виллерслев). Публикуем ее перевод на русский язык с предисловием Л.С.Клейна.
|
Анализ ДНК представителей минойской и микенской цивилизаций доказал их генетическое родство между собой, а также с современными греками. Показано, что основной вклад в формирование минойцев и микенцев внесли неолитические популяции Анатолии. Авторы обнаружили у них генетический компонент, происходящий с Кавказа и из Ирана, а у микенцев – небольшой след из Восточной Европы и Сибири.
|
Африка – прародина современного человека. Тем не менее генетические данные о древнем населении Африки до сего времени были совершенно незначительными – всего один прочитанный древний геном из Эфиопии возрастом 4,5 тысячи лет. Причины понятны – в экваториальном и тропическом климате ДНК плохо сохраняется и непригодна для изучения. Но вот сделан большой шаг вперед в этом направлении – секвенированы сразу семь древних африканских геномов, о чем поведала статья генетиков из Университета Упсалы, Швеция, опубликованная на сайте препринтов.
|
Публикуем заключительную часть статьи археологов из Одесского университета проф. С.В. Ивановой и к.и.н. Д.В. Киосака и археогенетика, проф. Grand Valley State University А.Г. Никитина. Предмет исследования — археологическая и культурная картина Северо-Западного Причерноморья эпохи энеолита — ранней бронзы и гипотеза о миграции населения ямной культуры в Центральную Европу.
|
Продолжаем публиковать статью археологов из Одесского университета проф. С.В. Ивановой и к.и.н. Д.В. Киосака и археогенетика, проф. Grand Valley State University А.Г. Никитина. Предмет исследования - археологическая и культурная картина Северо-Западного Причерноморья эпохи энеолита - ранней бронзы и гипотеза о миграции населения ямной культуры в Центральную Европу.
|
Представляем статью крупнейшего специалиста по степным культурам, проф. Одесского университета С.В. Ивановой, археолога из Одесского университета Д.В. Киосака и генетика, работающего в США, А.Г. Никитина. В статье представлена археологическая и культурная картина Северо-Западного Причерноморья эпохи энеолита - ранней бронзы и критический разбор гипотезы о миграции населения ямной культуры в Центральную Европу. Публикуем статью в трех частях.
|
Новые детали взаимоотношений современного человека с неандертальцами получены по анализу митохондри альной ДНК неандертальца из пещеры в Германии. Предложенный авторами сценар ий предполагает раннюю миграцию предков сапиенсов из Африки в Европу, где они метисировались с неандертальцами, оставив им в наследство свою мтДНК.
|
Изучив митохондриальную ДНК древних и современных армян, генетики делают вывод о генетической преемственности по материнским линиям наследования в популяциях Южного Кавказа в течение 8 тысяч лет. Многочисленные культурные перемены, происходящие за это время, не сопровождались изменениями в женской части генофонда.
|
Исследование генофонда парсов – зороастрийцев Индии и Пакистана – реконструировало их генетическую историю. Парсы оказались генетически близки к неолитическим иранцам, так как покинули Иран еще до исламизации. Несмотря на преимущественное заключение браков в своей среде, переселение в Индию оставило генетический след в популяции парсов. Оно сказалось в основном на их митохондриальном генофонде за счет ассимиляции местных женщин.
|
На прошедшем форуме «Ученые против мифов-4», организованном порталом «Антропогенез.ру», состоялась специальная конференция «Ученые против мифов-профи» - для популяризаторов науки. В профессиональной среде обсуждались способы, трудности и перспективы борьбы с лженаукой и популяризации науки истинной.
|
С разрешения авторов публикуем диалог д.и.н. Александра Григорьевича Козинцева и проф. Льва Самуиловича Клейна, состоявшийся в мае 2017 г.
|
С разрешения автора и издательства перепечатываем статью доктора историч. наук А.Г.Козинцева, опубликованную в сборнике, посвященном 90-летию Л.С.Клейна (Ex ungue leonem. Сборник статей к 90-летию Льва Самуиловича Клейна. СПб: Нестор-история, 2017. С.9-12).
|
Конференция «Позднепалеолитические памятники Восточной Европы», состоявшаяся в НИИ и Музее Антропологии МГУ, была посвящена 100-летию со дня рождения Марианны Давидовны Гвоздовер (1917-2004) – выдающегося археолога, специалиста по палеолиту. Участники конференции с большой теплотой вспоминали ее как своего учителя, а тематика докладов отражала развитие ее идей.
|
В журнале Science опубликованы размышления о роли исследований древней ДНК в представлениях об истории человечества и о непростых взаимодействиях генетиков с археологами. Одна из основных сложностей заключается в неоднозначных связях между популяциями и археологическими культурами. Решение сложных вопросов возможно только путем глубокой интеграции генетики, археологии и других наук.
|
По 367 митохондриальным геномам построено дерево гаплогруппы U7, определена ее прародина и описано распространение основных ветвей. Некоторые из них связывают с демографическими событиями неолита.
|
Казахские, российские и узбекские генетики исследовали генофонд населения исторического региона Центральной Азии – Трансоксианы по маркерам Y-хромосомы. Оказалось, что основную роль в структурировании генофонда Трансоксианы играет не географический ландшафт, а культура (хозяйственно-культурный тип): земледелие или же кочевое скотоводство. Показано, что культурная и демическая экспансии могут быть не взаимосвязаны: экспансия арабов не оказала значимого влияния на генофонд населения Трансоксианы, а демическая экспансия монголов не оказала значимого влияния на его культуру.
|
Российские антропологи исследовали особенности морфологии средней части лица в популяциях Северо-Восточной Европы в связи с факторами климата. Оказалось, что адаптации к низким температурам у них иные, чем у народов Северной Сибири. Полученные результаты помогут реконструировать адаптацию к климату Homo sapiens верхнего палеолита, так как верхнепалеолитический климат был более всего похож на современный климат Северо-Восточной Европы. Таким образом, современные северо-восточные европейцы могут послужить моделью для реконструкции процессов, происходивших десятки тысяч лет назад.
|
Немецкие генетики успешно секвенировали митохондриальную и проанализировали ядерную ДНК из египетских мумий разных исторических периодов. Они показали, что древние египтяне были генетически близки к ближневосточному населению. Современные египтяне довольно сильно отличаются от древних, главным образом долей африканского генетического компонента, приобретенного в поздние времена.
|
Данные по четырем древним геномам из бассейна Нижнего Дуная указали на долгое мирное сосуществование местных охотников-собирателей и мигрировавших земледельцев в этом регионе. На протяжении нескольких поколений между ними происходило генетическое смещение, а также передача культурных навыков.
|
Цвет кожи человека сформировался под сильным давлением естественного отбора и определяется балансом защиты от ультрафиолета и необходимого уровня синтеза витамина D. Цвет волос и радужной оболочки глаза, хотя в основном определяется тем же пигментом, в меньшей степени продукт естественного отбора и находится под большим влиянием других факторов. Одни и те же гены могут влиять на разные пигментные системы, а комбинация разных аллелей может давать один и тот же результат.
|
Юго-Восточная Европа в неолите служила местом интенсивных генетических и культурных контактов между мигрирующими земледельцами и местными охотниками-собирателями, показывает исследование 200 древних геномов из этого региона. Авторы описали разнообразие европейских охотников-собирателей; нашли, что не все популяции, принесшие земледелие в Европу, происходят из одного источника; оценили долю степного компонента в разных группах населения; продемонстрировали, что в смешении охотников-собирателей с земледельцами имел место гендерный дисбаланс – преобладание мужского вклада от первых.
|
Культурная традиция колоковидных кубков (одна из самых широко распространенных культур в позднем неолите/бронзовом веке), по-видимому, распространялась по Европе двумя способами – как передачей культурных навыков, так и миграциями населения. Это выяснили палеогенетики, представив новые данные по 170 древним геномам из разных регионов Европы. В частности, миграции с континентальной Европы сыграли ведущую роль в распространении ККК на Британские острова, что привело к замене 90% генофонда прежнего неолитического населения.
|
Российские антропологи провели новое исследование останков человека с верхнепалеолитической стоянки Костёнки-14 с использованием современных статистических методов анализа. Они пришли к выводу о его принадлежности к европеоидному типу и отсутствии австрало-меланезийских черт в строении черепа и зубной системы. Примечательно, что этот вывод согласуется с данными палеогенетиков.
|
Профессор Тоомас Кивисилд, один из ведущих геномных специалистов, представляющий Кембриджский университет и Эстонский биоцентр, опубликовал обзор по исследованиям Y-хромосомы из древних геномов. В этой обобщающей работе он сфокусировался на данных по Y-хромосомному разнообразию древних популяций в разных регионах Северной Евразии и Америки.
|
С разрешения редакции публикуем статью д.и.н. О.В.Шарова (Институт истории материальной культуры РАН) о роли выдающегося археолога д.и.н. М. Б. Щукина в решении проблемы природы черняховской культуры. В следующих публикациях на сайте можно будет познакомиться непосредственно с трудами М. Б. Щукина.
|
Перепечатываем статью выдающегося археолога М.Б.Щукина «Рождение славян», опубликованную в 1997 г. в сборнике СТРАТУМ: СТРУКТУРЫ И КАТАСТРОФЫ. Сборник символической индоевропейской истории. СПб: Нестор, 1997. 268 с.
|
Ученым удалось выделить древнюю мтДНК, в том числе неандертальцев и денисовцев, из осадочных отложений в пещерах, где не сохранилось самих костей. Авторы считают, что этот способ может значительно увеличить количество древних геномов.
|
Авторы находки в Южной Калифорнии считают, что метки на костях мастодонта и расположение самих костей говорят о следах человеческой деятельности. Датировка костей показала время 130 тысяч лет назад. Могли ли быть люди в Северной Америке в это время? Кто и откуда? Возникают вопросы, на которые нет ответов.
|
Представляем обзор статьи британского археолога Фолкера Хейда с критическим осмыслением последних работ палеогенетиков с археологических позиций.
|
Публикуем полную печатную версию видеоинтревью, которое несколько месяцев назад Лев Самуилович Клейн дал для портала "Русский материалист".
|
И снова о ямниках. Археолог Кристиан Кристиансен о роли степной ямной миграции в формировании культуры шнуровой керамики в Европе. Предлагаемый сценарий: миграция мужчин ямной культуры в Европу, которые брали в жены местных женщин из неолитических общин и формировали культуру шнуровой керамики, перенимая от женщин традицию изготовления керамики и обогащая протоиндоевропейский язык земледельческой лексикой.
|
Анализ древней ДНК из Эстонии показал, что переход от охоты-рыболовства-собирательства к сельскому хозяйству в этом регионе был связан с прибытием нового населения. Однако основной вклад внесла не миграция неолитических земледельцев из Анатолии (как в Центральной Европе), а миграция бронзового века из степей. Авторы пришли к выводу, что степной генетический вклад был, преимущественно, мужским, а вклад земледельцев Анатолии – женским.
|
Российские генетики изучили по Y-хромосоме генофонд четырех популяций коренного русского населения Ярославской области. Результаты указали на финно-угорский генетический след, но вклад его невелик. Наиболее ярко он проявился в генофонде потомков жителей города Молога, затопленного Рыбинским водохранилищем, что подтверждает давнюю гипотезу об их происхождении от летописных мерян. В остальных популяциях финно-угорский генетический пласт был почти полностью замещен славянским. Причем результаты позволяют выдвинуть гипотезу, что славянская колонизация шла преимущественно по «низовому» ростово-суздальскому пути, а не по «верховому» новгородскому.
|
Публикуем официальный отзыв д.ф.н. и д.и.н., проф. С.П.Щавелева на диссертацию и автореферат диссертации И.П. Лобанковой «Пассионарность в динамике культуры: философско-методологическая реконструкция культуры протогорода Аркаим», представленной на соискание ученой степени доктора философских наук.
|
В коротком сообщении, появившемся на сайте препринтов, его авторы – Иосиф Лазаридис и Дэвид Райх (Медицинская школа Гарварда), опровергают вывод, опубликованный недавно в статье Goldberg et al., о которой мы писали на сайте.
|
Продолжаем ответ на "этнический портрет среднестатистического россиянина" от компании "Генотек" . Часть третья, от специалиста по генетической генеалогии и блогера Сергея Козлова.
|
Продолжаем публиковать ответ на "этнический портрет среднестатистического россиянина" от компании "Генотек" . Часть вторая, от генетика, д. б. н., профессора Е.В.Балановской.
|
Публикуем наш ответ на опубликованный в массовой печати "этнический портрет среднестатистического россиянина" от компании "Генотек" . Часть первая.
|
Размещаем на сайте препринт статьи, предназначенной для Acta Archaeologica (Kopenhagen), для тома, посвященного памяти выдающегося датского археолога Клауса Рандсборга (1944 – 2016), где она будет опубликована на английском языке.
|
Известнейший российский археолог Лев Клейн написал две новые книги. Как не потерять вдохновение в работе над книгой? Когда случилось ограбление века? И что читать, если хочешь разбираться в археологии? Лев Самуилович отвечает на вопросы корреспондента АНТРОПОГЕНЕЗ.РУ
|
Публикуем комментарий проф. Л.С.Клейна на докторскую диссертацию И.П. Лобанковой «Пассионарность в динамике культуры: Философско-методологическая реконструкция культуры протогорода Аркаим».
|
Российские генетики исследовали генофонд народов Передней Азии и нашли интересную закономерность: наиболее генетически контрастны народы, живущие в горах и на равнине. Оказалось, что большинство армянских диаспор сохраняет генофонд исходной популяции на Армянском нагорье. По данным полного секвенирования 11 Y-хромосом авторы построили филогенетическое дерево гаплогруппы R1b и обнаружили на этом дереве помимо известной западноевропейской новую восточноевропейскую ветвь. Именно на ней разместились варианты Y-хромосом степных кочевников ямной культуры бронзового века. А значит, не они принести эту мужскую линию в Западную Европу.
|
В издательстве ЕВРАЗИЯ в Санкт-Петербурге вышла научно-популярная книга проф. Льва Самуиловича Клейна "Первый век: сокровища сарматских курганов". Она посвящена двум самым выдающимся памятникам сарматской эпохи нашей страны — Новочеркасскому кладу (курган Хохлач) и Садовому кургану.
|
Исследуя останки из захоронений степных кочевников железного века – скифов – методами краниометрии (измерение параметров черепов) и методами анализа древней ДНК, антропологи и генетики пришли к сопоставимым результатам. Те и другие специалисты обнаруживают близость кочевников культуры скифов к культурам кочевников бронзового века Восточной Европы. Антропологическими и генетическими методами у носителей скифской культуры выявляется также центральноазиатский (антропологи) либо восточноазиатско-сибирский (генетики) вклад. Что касается прародины скифов – европейские или азиатские степи – то по этому вопросу специалисты пока не пришли к единому мнению.
|
Древняя ДНК может рассказать не только о миграциях и демографической истории наших предков, но и о социальном устройстве общества. Пример такого исследования – работа генетиков из Университета Пенсильвании, опубликованная в журнале Nature Communication.
|
Представляем сводку археологических культур, представленных на страницах Словарика. Пока - список по алфавиту.
|
Публикуем статью Сергея Козлова с результатами анализа генофондов некоторых северных народов в свете данных из монографии В.В.Напольских "Очерки по этнической истории".
|
Анализ митохондриальной ДНК представителей трипольской культуры Украины показал ее генетическое происхождение по материнским линиям от неолитических земледельцев Анатолии с небольшой примесью охотников-собирателей верхнего палеолита. Популяция трипольской культуры из пещеры Вертеба генетически сходна с другими популяциями европейских земледельцев, но более всего – с популяциями культуры воронковидных кубков.
|
Анализ древней ДНК мезолита и неолита Балтики и Украины не выявил следов миграции земледельцев Анатолии, аналогичный найденным в неолите Центральной Европы. Авторы работы предполагают генетическую преемственность от мезолита к неолиту в обоих регионах. Они также нашли признаки внешнего влияния на генофонд позднего неолита, наиболее вероятно, это вклад миграции из причерноморских степей или из Северной Евразии. Определенно, неолит как в регионе Балтики, так и на Днепровских порогах (Украина) развивался иными темпами, чем в Центральной и Западной Европе, и не сопровождался такими масштабными генетическими изменениями.
|
Рассказ о генетико-антропологической экспедиции Медико-генетического научного центра и Института общей генетики РАН, проведенной в конце 2016 года в Тверскую область для исследования генофонда и создания антропологического портрета тверских карел и тверских русских.
|
Изучив митохондриальную ДНК из погребений энеолита и бронзового века в курганах Северного Причерноморья, генетики сделали вывод о генетической связи популяций степных культур с европейскими мезолитическими охотниками-собирателями.
|
9 января исполнился год со дня скоропостижной смерти смерти археолога и этнографа Владимира Александровича Кореняко, ведущего научного сотрудника Государственного музея искусства народов Востока, одного из авторов нашего сайта. С разрешения издательства перепечатываем его статью об этнонационализме, которая год назад была опубликована в журнале "Историческая экспертиза" (издательство "Нестор-история").
|
1 февраля на Биологическом факультете МГУ прошло Торжественное заседание, посвященное 125-летию со дня рождения Александра Сергеевича Серебровского, русского и советского генетика, члена-корр. АН СССР, академика ВАСХНИЛ, основателя кафедры генетики в Московском университете.
|
В совместной работе популяционных генетиков и генетических генеалогов удалось построить филогенетическое дерево гаплогруппы Q3, картографировать распределение ее ветвей, предположить место ее прародины и модель эволюции, начиная с верхнего палеолита. Авторы проследили путь ветвей гаплогруппы Q3 от Западной и Южной Азии до Европы и конкретно до популяции евреев ашкенази. Они считают, что этот удачный опыт послужит основой для дальнейшего сотрудничества академической и гражданской науки.
|
В конце ноября прошлого года в Москве прошла Всероссийская научная конференция «Пути эволюционной географии», посвященная памяти профессора Андрея Алексеевича Величко, создателя научной школы эволюционной географии и палеоклиматологии. Конференция носила междисциплинарный характер, многие доклады были посвящены исследованию географических факторов расселения человека по планете, его адаптации к различным природным условиям, влиянию этих условий на характер поселений и пути миграции древнего человека. Представляем краткий обзор некоторых из этих междисциплинарных докладов.
|
Публикуем статью Сергея Козлова о структуре генофонда Русского Севера, написанную по результатам анализа полногеномных аутосомных данных, собранных по научным и коммерческим выборкам.
|
В журнале Science Advances опубликованы результаты исследования геномов двух индивидов из восточноазиатской популяции эпохи неолита. Определено их генетическое сходство с ныне живущими популяциями. До сих пор исследования древней ДНК очень мало затрагивали регион Восточной Азии. Новые данные были получены при исследовании ДНК из останков двух женщин, найденных в пещере «Чертовы ворота» в Приморье, их возраст составляет около 7700 лет. Эти индивиды принадлежали к популяции охотников-рыболовов-собирателей, без каких-либо признаков производящего хозяйства, хотя было показано, что из волокон диких растений они изготавливали текстиль.
|
Обзор истории заселения всего мира по данным последних исследований современной и древней ДНК от одного из самых известных коллективов палеогенетиков под руководством Эске Виллерслева. Представлена картина миграций в глобальном масштабе, пути освоения континентов и схемы генетических потоков между человеком современного типа и древними видами человека.
|
Изучение Y-хромосомных портретов крупнейшей родоплеменной группы казахов в сопоставлении с данными традиционной генеалогии позволяет выдвинуть гипотезу, что их генофонд восходит к наследию народов индоиранской языковой семьи с последующим генетическим вкладом тюркоязычных и монголоязычных народов. Вероятно, основным родоначальником большинства современных аргынов был золотоордынский эмир Караходжа (XIV в.) или его ближайшие предки.
|
Путем анализа Y-хромосомных и аутосомных данных современного населения Юго-Западной Азии генетики проследили пути, по которым шло заселение этой территории после окончания Последней ледниковой эпохи. Они выделили три климатических убежища (рефугиума), которые стали источником миграций в регионе, и определили время расхождения ветвей Y-хромосомы в популяциях. Полученные результаты авторы обсуждают в связи с археологическими данными и работами по древней ДНК.
|
Генетики секвенировали четыре генома Yersinia pestis эпохи бронзового века. Их сравнение с другими древними и современными геномами этой бактерии привело к гипотезе, что чума в Европе появилась со степной миграцией ямной культуры, а затем вернулась обратно в Центральную Азию.
|
Исследование показало, что подавляющее большинство американских антропологов не считают расы биологической реальностью, не видят в расовой классификации генетической основы и не считают, что расу нужно учитывать при диагностике и лечении заболеваний. Сравнение показало, что антропологов, не признающих расы, в 2013 году стало радикально больше, чем 40 лет назад. Cтатья с результатами этого исследования опубликована в American Journal of Physical Anthropology.
|
Отзыв проф. Л.С.Клейна о книге Д.В.Панченко «Гомер, „Илиада”, Троя», вышедшей в издательстве «Европейский Дом».
|
В конце уходящего 2016 года попробуем подвести его итоги – вспомнить самые интересные достижения на перекрестке наук, изучающих историю народонаселения – археологии, антропологии, генетики, палеогеографии, лингвистики и др. Конечно, наш взгляд субъективен, поскольку мы смотрим через окно сайта «Генофонд.рф», ориентируясь на опубликованные на нем материалы. По той же причине в научных итогах мы вынужденно делаем крен в генетику. Будем рады если эта картина станет полнее с помощью комментариев от наших читателей.
|
Коллектив генетиков и историков изучил генофонды пяти родовых объединений (кланов) северо-восточных башкир. Преобладание в их Y-хромосомных «генетических портретах» одного варианта гаплогрупп указывает на единый генетический источник их происхождения – генофонд прото-клана. Выдвинута гипотеза, что формирование генофонда северо-восточных башкир связано с трансуральским путем миграций из Западной Сибири в Приуралье, хорошо известном кочевникам в эпоху раннего железного века и средневековья.
|
Перепечатываем статью О.П.Балановского, опубликованную татарским интернет-изданием "Бизнес-онлайн" - ответ критикам исследования генофондов татар.
|
Изучение Y-хромосомных генофондов сибирских татар выявило генетическое своеобразие каждого из пяти субэтносов. По степени различий между пятью популяциями сибирские татары лидируют среди изученных коллективом народов Сибири и Центральной Азии. Результаты позволяют говорить о разных путях происхождения генофондов сибирских татар (по данным об отцовских линиях): в каждом субэтносе проявляется свой субстрат (вклад древнего населения) и свой суперстрат (влияние последующих миграций).
|
Дискуссия, вызванная статьей о генофонде татар в "Вестнике МГУ", вылилась на страницы интернет-издания "Бизнес-онлайн". Публикуем письмо, отправленное д.б.н., профессором РАН О.П. Балановским 17 декабря 2016 года одному из участников этой дискуссии, д.и.н., специалисту по этногенезу татарского народа И.Л.Измайлову. Письмо, к сожалению, осталось без ответа.
|
Исследование Y-хромосомы туркменской популяции в Каракалпакстане (на территории Узбекистана) выявило сильное доминирование гаплогруппыQ, что, вероятно, объясняется их преобладающей принадлежностью к одному роду (йомуд). По генетическим расстояниям туркмены Каракалпакстана оказались близки к географически далеким от них туркменам Ирана и Афганистана и далеки от своих географических соседей – узбеков и каракалпаков.
|
Генофонды популяций с этнонимом «татары» трех регионов Евразии - крымские, поволжские и сибирские – исследованы путем анализа Y-хромосомы. Этнотерриториальные группы татар оказались генетически очень разнообразны. В генофонде поволжских татар преобладают генетические варианты, характерные для Приуралья и Северной Европы; в генофонде крымских татар преобладает вклад переднеазиатского и средиземноморского населения; популяции сибирских татар наиболее разнообразны: одни включают значительный сибирский генетический компонент, в других преобладают генетические линии из юго-западных регионов Евразии.
|
Популяционно-генетическую историю друзов британский генетик Эран Элхаик исследует методом GPS (geographic population structure). Критика специалистов в адрес предыдущих работ с использованием данного метода, вызывает вопросы и к данной работе.
|
Опубликовано на сайте Антропогенез.ру
|
В пределах 265 языковых семей исследователи показали корреляцию между лексикой разных языков и географическим положением. На примере 11 популяций из Африки, Азии и Австралии выявили корреляцию лексических расстояний между популяциями с фенотипическими расстояниями, самую высокую – по строению лицевой части черепа. Делается вывод о том, что лингвистические показатели можно использовать для реконструкции недавней истории популяций, но не глубокой истории.
|
Представляяем обзор некоторых докладов на прошедшей в Москве конференции «Эволюционный континуум рода Homo», посвященной 125-летию со дня рождения выдающегося русского антрополога Виктора Валериановича Бунака (1891–1979), иными словами, на Бунаковских чтениях.
|
Из-за чего случился бронзовый коллапс, как исчезла знаменитая майкопская культура, в чём заблуждаются сторонники «новой хронологии» и какие байки живут среди археологов, порталу АНТРОПОГЕНЕЗ.РУ рассказал Александр Скаков - кандидат исторических наук, научный сотрудник Отдела бронзового века Института археологии РАН.
|
В Москве завершила свою работу международная антропологическая конференция, посвященная 125-летию выдающегося русского антрополога Виктора Валериановича Бунака. Приводим краткий обзор ее итогов, опубликованный на сайте Центра палеоэтнологических исследований.
|
К сожалению, эхо от казанского интервью академика Валерия Александровича Тишкова (директора Института этнологии и антропологии РАН) не затихло, а рождает все новые недоразумения, которые отчасти уже объяснены на нашем сайте. Чтобы приостановить снежный ком, нам все же придется дать разъяснения неточностей, его породивших.
|
Статья американских и шведских исследователей (Goldberg et al.), опубликованная на сайте препринтов, вновь обращается к дискуссионной проблеме миграций в эпоху неолита и бронзового века. В работе исследуется вопрос о доле мужского и женского населения в составе мигрирующих групп, которые сформировали генофонд Центральной Европы. Авторы проверяют исходную гипотезу, что миграции из Анатолии в раннем неолите и миграции из понто-каспийских степей в течение позднего неолита и бронзового века были преимущественно мужскими.
|
Специалист по этногенезу тюркских народов Жаксылык Сабитов комментирует миф о финно-угорском происхождении татар, который без всяких на то оснований приписывается генетикам.
|
О.П.Балановский о том, как проходило обсуждение доклада А.В.Дыбо «Происхождение и родственные связи языков народов России» на Президиуме РАН.
|
Публикуем изложение доклада чл-корр. РАН Анны Владимировны Дыбо (Институт языкознания РАН), размещенное на сайте РАН.
|
Полное секвенирование геномов 83 австралийских аборигенов и 25 жителей Папуа Новая Гвинея позволило исследователям реконструировать историю заселения этой части света в пространстве и во времени. Они подтвердили, что предки австралийских аборигенов и папуасов Новой Гвинеи очень рано отделились от предков материковой Евразии. На ключевой вопрос о том, сколько раз человечество выходило из Африки – один или два, авторы отвечают с осторожностью. Большая часть их аргументов склоняет чашу весов к модели одного выхода, однако тот вариант, что их могло быть два, исследователи не отвергают.
|
Прочитав с высокой степенью надежности 379 геномов из 125 популяций со всего мира, исследователи уточнили картину современного генетического разнообразия и пути древних миграций, которые к нему привели. В частности, в геномах папуасов Новой Гвинеи они нашли небольшой вклад ранней миграционной волны из Африки, которая не оставила следов в геномах материковой Евразии.
|
Полное секвенирование 300 геномов из 142 популяций со всего мира дало возможность исследователям добавить важные фрагменты в мозаику геномного разнообразия населения планеты. Они пересчитали вклад неандертальцев и денисовцев в современный геном в глобальном масштабе, вычислили, как давно разошлись между собой разные народы, оценили степень гетерозиготности в разных регионах. Наконец, авторы уточнили источник генофонда жителей Австралии и Новой Гвинеи, показав, что они происходят от тех же популяций, что и жители остальной Евразии.
|
Приводим экспертное мнение Жаксылыка Сабитова (Евразийский Национальный Университет, Астана), специалиста по истории Золотой орды и этногенезу тюркских народов, по недавно опубликованной в журнале PLоS ONE статье .
|
В журнале PLOS Genetics опубликованы результаты широкогеномного (в пределах всего генома) исследования ассоциаций (GWAS) различных черт лица. У 3118 жителей США европейского происхождения авторы провели трехмерное измерение 20 лицевых признаков и анализ однонуклеотидного полиморфизма (около 1 млн SNP). Обнаружили достоверную связь полиморфных участков генома с шириной черепа, шириной расстояния между внутренними углами глаз, шириной носа, длиной крыльев носа и глубиной верхней части лица.
|
Коллектив генетиков и биоинформатиков опубликовал обзор истории изучения древней ДНК, основных трудностей в ее изучении и методов их преодоления. Авторы представили новейшие знания о путях миграций и распространения населения, полученные путем анализа древних геномов, и показали, какую революционную роль анализ палеоДНК сыграл в популяционной и эволюционной генетике, археологии, палеоэпидемиологии и многих других науках.
|
Проект по секвенированию более 60 тысяч экзомов (часть генома, кодирующая белки) в популяциях на разных континентах выявил гены, устойчивые к мутированию, показал, сколько носимых нами мутаций полностью блокируют синтез белка, а также значительно приблизил специалистов к пониманию природы редких заболеваний.
|
Российские генетики определили полную последовательность шести митохондриальных геномов древних людей, обитавших на территории Северного Кавказа на рубеже неолита и бронзы.
|
Сравнив фенотипические расстояния между 10 популяциями по показателям формы черепа и генетические расстояния по 3 345 SNP, исследователи нашли корреляции между ними. Они утверждают, что форма черепа в целом и форма височных костей может быть использована для реконструкции истории человеческих популяций.
|
Изучен генофонд популяции польско-литовских татар (липок), проживающих в Белоруссии. В их генофонде примерно две трети составляет западноевразийский компонент и одну треть – восточноевразийский. Очевидно, последний отражает влияние дальних миграций – степных кочевников Золотой Орды, поселившихся в Центральной и Восточной Европе.
|
Лингвисты из Кембриджского и Оксфордского университетов, разработали технологию, которая, как они утверждают, позволяет реконструировать звуки праиндоевропейского языка. Сообщение об этом опубликовано на сайте Кембриджского университета http://www.cam.ac.uk/research/features/time-travelling-to-the-mother-tongue.
|
Перепечатываем статью Павла Флегонтова и Алексея Касьяна, опубликованную в газете "Троицкий вариант", с опровержением гипотезы английского генетика Эрана Элхаика о хазарском происхождении евреев ашкеназов и славянской природе языка идиш. Эта популярная статья вышла параллельно с научной статьей с участием этих же авторов в журнале Genome Biology and Evolution.
|
15 июля в Еженедельной газете научного сообщества "Поиск" опубликовано интервью с О.П. Балановским. Подробности по ссылке:
|
Турсервис Momondo сделал генетические тесты и записал реакцию на их результаты. Видео получилось простым и понятным. А что думает об этом популяционная генетика?
|
В только что опубликованной статье была подробно изучена история распространения одной из самых широко встречающихся в Евразии Y-хромосомных гаплогрупп – N. По данным полного секвенирования Y-хромосомы было построено филогенетическое дерево и описано подразделение гаплогруппы на ветви и субветви. Оказалось, что большинство из них имеют точную географическую но не лингвистическую привязку (встречаются в популяциях различных языковых семей).
|
Новое исследование генетических корней евреев ашкеназов подтвердило смешанное европейско-ближневосточное происхождение популяции. В составе европейского предкового компонента наиболее существенный генетический поток ашкеназы получили из Южной Европы.
|
Опубликована единственная на настоящий момент работа, посвященная исследованию генофонда верхнедонских казаков. Для изучения генофонда казаков использован новый инструмент - программа Haplomatch, позволяющая производить сравнение целых массивов гаплотипов. Удалось проследить, что формирование генофонда казаков верхнего Дона шло преимущественно за счет мигрантов из восточно-славянских популяций (в частности с южно-, центрально - русских и украинцев). Также обнаружено небольшое генетическое влияние ногайцев, вероятно вызванное их вхождением в Войско Донское в составе «татарской прослойки». Сходства с народами Кавказа у донских казаков не обнаружено.
|
Публикуем перевод статьи Душана Борича и Эмануэлы Кристиани, в которой рассматриваются социальные связи между группами собирателей палеолита и мезолита в Южной Европе (на Балканах и в Италии). Социальные связи прослеживаются в том числе путем исследования и сопоставления технологий изготовления орудий и украшений.
|
Используя традиционные подходы и свой собственный новый метод, специалисты изучили происхождение коренных народов Сибири. Для популяций Южной Сибири, они реконструировали последовательность генетических потоков, которые смешивались в генофонде.
|
Анализ древней ДНК с Ближнего Востока показал, что большой вклад в генофонд первых ближневосточных земледельцев внесла древняя линия базальных евразийцев; что в пределах Ближнего Востока популяции земледельцев генетически различались по регионам, и между охотниками-собирателями и первыми земледельцами в каждом регионе имелась генетическая преемственность.
|
Представляем обобщающую статью по культурам верхнего палеолита, которая может служить пояснением для соответствующих статей в Словарике, посвященных отдельным культурам верхнего палеолита.
|
Форум «Ученые против мифов», организованный порталом «Антропогенез.ру», прошел в Москве 5 июня. Организаторы обещают скоро выложить записи докладов. Пока же представляем основные тезисы, прозвучавшие в выступлениях участников форума.
|
Анализ древней и современной ДНК собак, включая полностью секвенированный древний геном неолитической собаки из Ирландии и 605 современных геномов, привел исследователей к гипотезе, что человек независимо одомашнил волка в Восточной Азии и в Европе. Затем палеолитическая европейская популяция собак была частично замещена восточноазиатскими собаками.
|
Митохондриальная ДНК человека возрастом 35 тыс. лет назад из пещеры в Румынии оказалась принадлежащей к африканской гаплогруппе U6. Из этого исследователи сделали вывод о евразийском происхождении этой гаплогруппы и о том, что она была принесена в Северную Африку путем верхнепалеолитической обратной миграции.
|
Археологи провели исследование загадочных конструкций в форме кольца из обломков сталагмитов в пещере Брюникель на юго-западе Франции. Особенности конструкций, следы огня на них и соседство с костями говори т об их рукотворном происхождении. Датировка - 176.5 тысяч лет назад – указала на ранних неандертальцев.
|
Cпециалисты нашли шесть генов, вариации в которых влияют на черты лица человека. Все они экспрессируются при эмбриональной закладке лицевой части черепа, влияя на дифференцировку клеток костной и хрящевой ткани. Больше всего генетические вариации связаны с параметрами носа.
|
С разрешения автора перепечатываем статью доктора истор. наук Виктора Александровича Шнирельмана "Междисциплинарный подход и этногенез", опубликованную в сборнике "Феномен междисциплинарности в отечественной этнологи" под ред Г. А. Комаровой, М.: ИЭА РАН, 2016. С. 258-284.
|
Исследование показало, что популяция Бене-Исраэль, живущая в Индии, имеет смешанное еврейско-индийское происхождение. Причем вклад евреев передался в основном по мужским линиям наследования (по Y-хромосоме), а вклад индийцев – по женским (по мтДНК). Время же возникновения популяции оказалось не столь давним, как в легендах.
|
Пещера Шове известна во всем мире наскальными рисунками эпохи палеолита. Древние художники использовали ее для своего творчества в два этапа с перерывом. Причем один из этих этапов перекрывался по времени с периодом обитания здесь пещерных медведей. Авторы нового исследования реконструировали историю обитания пещеры, используя многочисленные датировки и моделирование.
|
История генофонда Европы до неолитизации очень мало изучена. Новое исследование под руководством трех лидеров в области древней ДНК приоткрывает дверь в события более далекого прошлого. Авторы проанализировали 51 образец древней ДНК и частично реконструировали картину движения популяций до и после Последнего ледникового максимума. Они попытались связать обнаруженные ими генетические кластеры, объединяющие древних индивидов в пространстве и во времени, с определенными археологическими культурами.
|
Новый метод молекулярно-генетической датировки, предложенный в статье команды Дэвида Райха, основан на сравнении древних и современных геномов по длине неандертальских фрагментов ДНК. В отличие от радиоуглеродной датировки, этот метод точнее работает на более старых образцах. С его помощью авторы также вычислили длину поколения (26-30 лет), предположив, что она существенно не менялась за 45 тысячелетий.
|
По рекордному на сегодняшний день количеству полностью секвенированных Y-хромосом (1244 из базы проекта «1000 геномов») исследователи построили новое разветвленное Y-хромосомное дерево и попытались связать экспансию отдельных гаплогрупп с историческими сведениями и археологическими данными.
|
Палитра геномных исследований в России разнообразна. Создаются генетические биобанки, исследуется генетическое разнообразие популяций, в том числе генетические варианты, связанные с заболеваниями в разных популяциях; российские специалисты вовлечены в полногеномные исследования, и на карте мира постепенно появляются секвенированные геномы из России.
|
Исследователи секвенировали геномы из Меланезии и нашли у них наибольшую долю включений ДНК древних видов человека, причем как неандертальского, так и денисовского происхождения. Новые данные позволили нарисовать уточненную картину генетических потоков между разными видами Homo.
|
С разрешения автора публикуем тезисы его доклада на предстоящей конференции в Томске.
|
Представляем перевод статьи североирландского и американского археолога, специалиста по индоеропейской проблематике, профессора Джеймса Патрика Мэллори. Эта статья представляет собою обобщающий комментарий к некоторым докладам на семинаре «Прародина индоевропейцев и миграции: лингвистика, археология и ДНК» (Москва, 12 сентября 2012 года).
|
Исследователи из Стэнфордского университета, проанализировав Y-хромосому неандертальцев, убедились в том, что в Y-хромосоме современного человека нет неандертальских фрагментов ДНК, в отличие от остальной части генома. Этому факту они постарались дать объяснение. Скорее всего, дело в антигенах гистосовместимости, которые препятствовали рождению мальчиков с неандертальскими генами в Y-хромосоме.
|
Исследовав 92 образца древней мтДНК коренных американцев, генетики реконструировали основные этапы заселения Америки, уточнив пути основных миграций и их время. Они также пришли к выводу о драматическом влиянии европейской колонизации на генетическое разнообразие коренного населения Америки.
|
Публикуем перевод критической статьи известного болгарского археолога Лолиты Николовой. Ее критика направлена на авторов одной из самой яркой статьи прошлого года «Massive migration from the steppes was a source for Indo-European Languages in Europe» (Haak et al., 2015), в которой авторы представляют свою гипотезу распространения индоевропейских языков в Европе.
|
Публикуем статью украинского археолога, доктора ист. наук, проф. Леонида Львовича Зализняка, специально переведенную им на русский язык для нашего сайта. Статья представляет собой критический анализ взглядов на происхождение индоевропейцев с позиций археологии и других наук.
|
Перепечатываем статью швейцарского лингвиста Патрика Серио, перевод которой был опубликован в журнале «Политическая лингвистика». В статье анализируется явление «Новой парадигмы» в области лингвистики в странах Восточной Европы. С точки зрения автора, это явление подходит под определение ресентимента.
|
Человек (Homo sapiens) – единственное в природе существо, которое может переносить из сознания на внешние носители фигуративные образы. В эволюции нет ничего, что бы предшествовало этой способности. Таким же уникальным свойством является способность к членораздельной речи, к языку. Звуковые сигналы в мире других живых существ заданы генетически. Возникает предположение, что эти две способности связаны между собой больше, чем нам кажется.
|
Генетический анализ популяции кетов – коренного народа Сибири, в сравнении с окружающим народами в бассейне Енисея выявил их наиболее тесную связь с карасукской культурой бронзового века Южной Сибири - именно в этом регионе находится гипотетическая прародина енисейской семьи языков. Более глубокие корни кетов уходят к ветви древних северных евразийцев. По опубликованным ранее и по новым данным, 5000-6000 лет назад генетический поток протянулся от сибирских популяций до культуры саккак (палеоэскимосов американской Арктики), и от саккак к носителям языков на-дене. Примечательно, что данная миграция согласуется с гипотезой о родстве енисейских языков и языков на-дене.
|
История взаимоотношений человека современного вида и неандертальцев оказалась непростой и долгой. Не только неандертальцы оставили след в нашем геноме. Обнаружен генетический поток и от Homo sapiens к предкам алтайских неандертальцев. Он указывает на раннюю - около 100 тысяч лет назад - метисацию, что происходила еще до основной волны выхода наших предков из Африки.
|
Статья является реакцией на публикацию коллектива американских авторов, отрицающих существование рас у человека и, более того, призывающих отменить и запретить использование самого термина «раса». Авторы обнаруживают полное незнание предмета обсуждения и научной литературы по проблеме расы. «Антирасовая кампания», уже давно развязанная в США и перекинувщаяся в научные центры Западной Европы, отнюдь не служит делу борьбы с расизмом, а наоборот, способствует появлению разного рода действительно расистских публикации, в том числе, в самих США. А методы проведения этой кампании напоминают времена лысенковщины в СССР.
|
Публикуем статью генетика д.б.н. Е.В. Балановской (вернее, раздел в сборнике «Проблема расы в российской физической антропологии» [М., Институт этнологии и антропологии РАН, 2002]). Сегодня эта статья, к сожалению, не менее актуальна, чем пятнадцать лет назад: недавно Science опубликовал статью с предложением отказаться от понятия «раса» в генетических исследованиях. И это при том, что именно генетические исследования доказывают реальность существования рас.
|
Авторы статьи в Science утверждают, что в современной генетике понятие «раса» - бесполезный инструмент при характеристике генетического разнообразия человечества. Учитывая проблемы, связанные с неправильным употреблением термина, они предлагают вообще от него отказаться. Правда, рассуждения авторов касаются только генетики, они не рассматривают понятие "раса" в рамках антропологии.
|
Генетики исследовали популяцию уйгуров, по одной из версий являющихся генетическими потомками тохаров. Через ареал уйгуров проходил Великий Шелковый путь, соединявший Восточную Азию с Центральной Азией и Европой. Результаты, полученные по STR маркерам Y-хромосомы, подтверждают гипотезу, что в формировании современного генофонда уйгуров сыграли почти равную роль как европейские так и восточноазиатские популяции, но все же с преобладанием вклада генофондов Западной Евразии.
|
Секвенирование 55 древних митохондриальных геномов (возраст – от 35 до 7 тысяч лет), выявило в них варианты, которые не встречены в современном населении Европы. Описав демографические изменения в их связи с изменениями климата, коллектив Йоханеса Краузе (Йена) пришел к выводу, что около 14,5 тысяч лет назад в Европе радикально изменился генофонд охотников-собирателей.
|
Евразийский вклад в генофонд африканских популяций существует, но не столь велик – он обнаруживается не на всем континенте, а в основном в Восточной Африке. Важно, что ошибка признана авторами статьи публично и бесконфликтно - это – признак «здоровья» генетического консорциума.
|
Публикуем статью проф. Л.С.Клейна (вышедшую в журнале "Археологические Вести", 21, 2015) о том, как д.х.н. А.А.Клесов, занявшись темой происхождения славян, связывает ее с вопросом о «норманнской теории», хотя это совсем другая тема - происхождения государственности у восточных славян.
|
Путем секвенирования геномов из семи популяций исследователи подтвердили картину расселения человека по континентам после выхода из Африки. Серия миграций сопровождалась снижением генетического разнообразия. По этой же причине с увеличением расстояния от Африки возрастает мутационный груз в популяциях.
|
Две статьи с данными по секвенированным древним геномам дополнили представления о том, какую роль играли исторические миграции – римского времени и англосаксонская – в формировании современного генофонда Великобритании. Так, уточненный генетический вклад англосаксонских переселенцев составляет около 40% в восточной Англии и 30% - в Уэльсе и Шотландии.
|
Четыре секвенированных генома древних жителей Ирландии (один эпохи неолита, три – бронзового века) указывают, что генофонд Британских островов, как и остальной Европы, сформировался при смешении западно-европейских охотников-собирателей с неолитическими земледельцами, прибывшими с Ближнего Востока, и с более поздней миграцией, берущей начало из степей Евразии.
|
11-13 октября в Йене, Германия в Институте наук об истории человека общества Макса Планка (Max Planck Institute for the Science of Human History) прошла первая междисциплинарная конференция, посвященная недавним генетическим открытиям о миграциях индоевропейцев. Генетики, археологи и лингвисты собрались вместе, чтобы обсудить, как полученные ими последние данные интегрируются в индоевропейскую проблему. Приводим обзор основных идей участников конференции, которые они изложили в своих выступлениях.
|
Публикуем рецензию д.и.н. профессора Ф.Х. Гутнова на брошюру г-на Тахира Моллаева (работника Национального парка «Приэльбрусье», бывшего заочника-филолога КБГУ), «Новый взгляд на историю осетинского народа». Редакция особо отмечает, что пантюркистская тенденция никогда в нашей науке не имела ни авторитета, ни поддержки..
|
Якутские лошади – самые северные на планете и самые морозоустойчивые. Прочитав два древних и девять современных геномов и использовав базу данных по другим геномам, команда российских и зарубежных исследователей нашла ответы на два вопроса. Первый вопрос - от каких древних популяций произошли современные якутские лошади. А второй – как им удалось приспособиться к экстремальным условиям якутского климата за такое короткое время.
|
Почти рождественская история с пропавшим листком, поиском автора и ответами проф. Л.С.Клейна на вопросы антинорманиста.
|
Провожая уходящий год, мы решили подвести итоги и выделить наиболее интересные, на наш взгляд, междисциплинарные исследования в области истории популяций, формирования генетического ландшафта мира и этногенеза, которые были опубликованы в 2015 году. Почти все они нашли свое отражение в материалах нашего сайта. Основные открытия года можно сгруппировать в несколько блоков.
|
Генетики исследовали варианты Y-хромосомы у 657 австралийских аборигенов. Среди них оказалось 56% носителей пришлых евразийских гаплогрупп и только 44% носителей коренных гаплогрупп. Авторы подтвердили гипотезу раннего (около 50 тыс. лет назад) заселения Австралии и длительной изоляции Австралии и Новой Гвинеи. Не найдено доказательств миграций в Австралию из Индии в голоцене. А вот европейская колонизация в конце XVIII века драматически снизила разнообразие коренных австралийских гаплогрупп.
|
Продолжаем публиковать фрагмент из книги О.П.Балановского "Генофонд Европы", посвященный анализу полногеномных маркеров ДНК - самых современных и наиболее информативных для анализа генофонда. В этой части описан метод анализа предковых компонентов и его отображение на геногеографических картах народов Европы
|
Следующий фрагмент книги О.П.Балановского "Генофонд Европы" посвящен полногеномным и широкогеномным маркерам ДНК. Это самые современные и наиболее информативные методы анализа генофонда. В первой части главы показано, как выявляемая с их помощью генетическая карта Европы соотносится с географической картой.
|
Продолжаем публиковать фрагмент из книги О.П.Балановского «Генофонд Европы», посвященный митохондриальной ДНК. В нем разбирается географическая и лингвистическая структурированность генофонда Европы, а также гаплотипическое разнообразие по мтДНК и эколого-генетический мониторинг.
|
Доклад доктора биол. наук Л.А.Животовского об изданной им книге «Неизвестный Лысенко» собрал аншлаг в Институте океанологии РАН. Собственно, не сам доклад, а последующее за ним обсуждение этой попытки реабилитации самой одиозной фигуры советской биологии. Свое мнение высказали и специалисты ненавидимой им генетики, и те, для которых драматические события, связанные с «народным академиком» прошлись по судьбам их семей.
|
В публикуемом фрагменте из книги О.П.Балановского «Генофонд Европы» речь идет об одной из трех систем для оценки геномного разнообразия – митохондриальной ДНК (мтДНК). Дается обзор изменчивости генофонда Европы по мтДНК и рассматриваются генетические взаимоотношения популяций в этом зеркале.
|
В статье обсуждается этимология названия города Суздаль, а также предлагается и обосновывается гипотеза происхождения ойконима Суздаль от реконструируемого гидронима Суздаль (Суздаля).
|
В новой статье команды Сванте Паабо представлены антропологические и генетические данные по двум образцам – двум зубам из Денисовой пещеры. Поскольку генетически подтвердилась их принадлежность к денисовскому человеку, а не к неандертальцам, число проанализированных геномов денисовцев теперь увеличилось до трех.
|
В докладе доктора филолог. наук О.А.Мудрака «Язык и тексты восточно-европейской руники» была представлена расшифровка и перевод рунических надписей памятников, найденных на территории Восточной Европы – от Днепра и Кавказа до Поволжья. Прочтение этих надписей привело к неожиданным заключениям относительно языка бытового и официального письма живших на этой территории народов. Почти все они оказались написаны на осетинском языке и очень немногие - на чечено-ингушском.
|
Масштабный научный проект по изучению генофонда (экзомов) коренного населения народов Урало-Поволжья, в том числе генофонда татар, поддержал экс-президент Минтимер Шаймиев. Проект вызвал шумиху среди татарских националистов и тех, кто приписывает ученым националистически ориентированные цели.
|
Последняя часть главы по древней ДНК из книги О.П.Балановского "Генофонд Европы" посвящена Европе бронзового века. Анализируя палеоДНК, генетики подтверждают представления археологов, что бронзовый век был временем активных миграций и радикальных изменений образа жизни. Все большее количество древних геномов позволяет реконструировать направления миграций и связать генетические потоки с конкретными археологическими культурами.
|
Этот фрагмент из главы о древней ДНК книги О.П.Балановского "Генофонд Европы" рассказывает о том, как с помощью изучения палеоДНК можно реконструировать очень важные процессы неолитизации Европы. В том числе, выяснить, какие древние популяции внесли вклад в формирование генофонда европейцев.
|
В следующем разделе главы о древней ДНК из книги О.П.Балановского "Генофонд Европы" мы узнаем о генетических исследованиях находок времен верхнего палеолита и мезолита на территории Евразии.
|
Очередной фрагмент из книги О.П.Балановского "Генофонд Европы" посвящен анализу древней ДНК. Охарактеризованы проблемы и перспективы направления, сложности лабораторной работы и наиболее успешные исследовательские коллективы. Обзор конкретных исследований начинается со среднего палеолита - с результатов анализа ДНК неандертальцев и денисовцев.
|
Секвенировав три древних генома (верхний палеолит и мезолит) из Грузии и Швейцарии, генетики предполагают, что популяция кавказских охотников-собирателей могла быть четвертым источником европейского генофонда. А ее генетический вклад был передан в Европу, Южную и Центральную Азию через миграции степной ямной культуры.
|
Публикуем отрывок из готовящейся к изданию книги проф. Л.С. Клейна "Хохлач и Садовый". В этом фрагменте разбирается вопрос об этнической принадлежности тех, кто оставил донские курганы. Исследователи высказывают разные предположения о том, кому принадлежали курганы: сарматам, аланам или аорсам. Автор останавливается и на том, кто такие аланы и почему разные народы стремятся приписать себе происхождение от них.
|
В этом разделе из книги О.П.Балановского "Генофонд Европы" описывается структура генофонда Европы в зависимости от двух факторов - географического положения и лингвистики. Европейские популяции объединяются в кластеры как по географическому, так и по лингвистическому принципу. Анализ этой структурированности дается на двух уровнях: межэтническом и внутриэтническом.
|
Публикуем очередной фрагмент из книги О.П.Балановского "Генофонд Европы" (выйдет в декабре 2015 г.). В нем представлен обобщенный анализ генофонда Европы по всем гаплогруппам на трех уровнях: региональном, этническом и субэтническом.
|
Публикуем вторую часть беседы с генетиком, специалистом по древней ДНК Вольфгангом Хааком (Max Planck Institute for the Science of Human History) на конференции в Санкт-Петербурге. Во второй части В.Хаак рассказывает Надежде Маркиной о роли, которая играет исследование древней ДНК в реконструкции истории популяций, и о важности мультидисциплинарного подхода.
|
Публикуем первую часть беседы с генетиком, специалистом по древней ДНК Вольфгангом Хааком (Max Planck Institute for the Science of Human History), которая состоялась в Санкт-Петербурге. В первой части Л.С.Клейн и В. Хаак говорят о том, как по изучению древней ДНК специалисты предположили вклад древнего населения степей в европейский генофонд и с какими культурами они его связывают.
|
В бронзовом веке чума была вполне обычным явлением, хотя в то время чумная бацилла еще не научилась передаваться с блохами и не могла вызывать самую опасную разновидность болезни – бубонную чуму. Время возникновения Yersinia pestis и ее этапы на пути превращения в возбудителя смертельной болезни – все это ученые выяснили, прочитав геномы бактерий из древних останков человека.
|
Публикуем следующий фрагмент из книги О.П.Балановского "Генофонд Европы" . В нем представлены карты всех гаплогрупп Y-хромосомы, по которым есть надежные данные об их распространении в Европе. Этот фрагмент можно рассматривать как первую версию Атласа Y-хромосомы в Европе.
|
Публикуем статью С.В.Кончи, посвященную описанию снега и прочих зимних атрибутов в общеиндоевропейском лексическом фонде. Многие специалисты трактуют «зимнию» лексику как указание на расположение прародины индоевропейцев.
|
Вышел новый номер журнала Stratum plus, посвященный раннеславянской археологии Подунавья «Славяне на Дунае. Обретение Родины» . Его редакторы реализовали грандиозный замысел – собрали в номере почти всех наиболее крупных специалистов в этой области, выступивших с обзорными статьями.
|
Последняя серия карт генетических расстояний (из книги О. Балановского «Генофонд Европы») от народов, ничем друг на друга не похожих – ни языком, ни географией. Но зато эти три генофонда окаймляют пространство народов, рассмотренных в пяти предыдущих сериях, и позволяют увидеть, насколько велики различия генофондов европейской окраины Евразии. Эти три этноса – албанцы, шведы, ногайцы - не только географически «расставлены» по трем «концам земли», но и генетически полярно различны, показывая масштаб разнообразия генофонда Европы.
|
В пятой серии карт (из книги О. Балановского «Генофонд Европы») мы видим степень близости к каждой из популяций Европы южных славян - македонцев, сербов, хорватов, боснийцев и герцеговинцев. Географически их объединяет принадлежность к Балканам, а генетическое своеобразие связывается с сохранением субстратного генофонда тех балканских племен и народов, которые стали говорить на славянских языках.
|
Публикуем четвертую серию карт генетических расстояний на основе гаплогрупп Y-хромосомы из книги О.П. Балановского «Генофонд Европы». Эти карты отражают генетический ландшафт северной окраины Балкан, где проживают разноязыкие народы, говорящие на языках трех лингвистических семей.
|
Эта серия карт очередного фрагмента из книги О.П. Балановского «Генофонд Европы» описывает разнообразие Y-хромосомного генофонда Волжско-Уральского региона. Рассмотрена только полоса соседствующих популяций - Башкортостана, Татарстана, Чувашии и Мордовии. Но несмотря на их относительно небольшой суммарный ареал, генофонды оказались своеобразны и даже загадочны.
|
Следующий фрагмент из книги О.П. Балановского «Генофонд Европы» описывает своеобразие генофондов западных и восточных славян. Карты генетических расстояний обобщают разнообразие гаплогрупп Y-хромосомы и позволяют самим убедиться, насколько каждая точка в ареале Европы генетически близка к средним параметрам каждого из народов западных и восточных славян: их генофонды оказались настолько близки, что им хочется дать имя "генофонд северных славян".
|
Публикуем фрагмент из книги О.П. Балановского "Генофонд Европы" (выйдет в декабре 2015 г.). Карты генетических расстояний позволят своими глазами увидеть, насколько генофонд отдельного народа похож на все остальные генофонды Европы. Представлены карты первой из шести серий - "Народы Северо-Восточной Европы": от карел и вепсов, от эстонцев и коми, от литовцев и латышей, от северных русских и финнов.
|
Экспертное мнение проф. Л.С.Клейна на статью С.А.Григорьева "Еще раз о концепции Т.В.Гамкрелидзе и В.В.Иванова и о критических этюдах в индоевропеистике".
|
Представлены итоги проекта «1000 геномов». Секвенированы геномы и экзомы для 2504 индивидов из 26 популяций пяти регионов. Описано свыше 88 млн генетических вариаций. Создана модель реконструкции демографической истории популяций и найдены новые мишени естественного отбора.
|
Замечания проф. Л.С.Клейна, высказанные с позиций археолога, относительно изложения материала по древним геномам в новой статье команды Райха. С точки зрения эксперта в статье недостаточно внимания уделено принадлежности изучаемых образцов конкретным археологическим культурам.
|
В дополненной статье команды Дэвида Райха про исследование естественного отбора по древней ДНК более чем вдвое увеличилось число проанализированных древних геномов. В результате авторы пришли к новым выводам относительно генетического родства популяций, носителей основных археологических культур от раннего неолита до поздней бронзы.
|
Публикуем раздел книги О.П. Балановского "Генофонд Европы" (выйдет из печати в декабре 2015 г.), посвященный чрезвычайно важному в изучении истории народов вопросу - датировках миграций и других исторических событий. Автор описывает способы, которым решают его популяционные генетики, генетические генеалоги, а также останавливается на подходах "ДНК-генеалогии" А.А. Клесова, разъясняя их ошибочность и лженаучность.
|
В заметке описывается проект Лаборатории востоковедения и сравнительно-исторического языкознания Школы актуальных гуманитарных исследований РАНХиГС, связанный с формализацией генетической классификации языков.
|
Захоронение предполагаемых останков цесаревича Алексея и великой княжны Марии Романовых - детей императора Николая II, отложено на неопределенное время. Поэтому предлагаем вновь открыть страницы непростой истории генетической идентификации костных останков из двух захоронений близ Екатеринбурга – именно эти генетические исследования убедили ученых в их принадлежности членам царской семьи. Это отражено в заключении межведомственной правительственной комиссии, но уголовное дело вновь открыто: предстоит повторная экспертиза. В ее преддверии итоги уже пройденного пути подвел директор Института общей генетики РАН член-корреспондент РАН Н.К. Янковский.
|
В статье дается краткая характеристика текущего состояния и актуальных проблем т. н. "ностратической" гипотезы, разработанной в 1960-е гг. В. М. Иллич-Свитычем и А. Б. Долгопольским и предполагающей дальнее генетическое родство между собой ряда крупных языковых семей Старого Света (как минимум - индоевропейской, уральской, алтайской, картвельской и дравидийской).
|
Впервые генетики секвенировали хорошо сохранившуюся в пещере древнюю ДНК с территории Африки, получив первый эталонный африканский геном. Сравнение этого генома с современными указал на масштаб евразийской обратной миграции в Африку, вклад которой составляет 4-7% в современных африканских геномах на всем континенте.
|
В Америке вышла книга британского философа Стивена Лича «Российские перспективы теоретической археологии. Жизнь и труд Льва С. Клейна». Клейна считают самым известным из современных российских археологов на Западе, его больше других переводили, но на деле знают о нем и его идеях очень мало.
|
На рабочем совещании по проекту "Российские геномы" присутствовали организаторы проекта и лидеры всех основных популяционно-генетических коллективов России. Предлагаем Вашему вниманию доклад О.П. Балановского, представленный на этой конференции. В нем, в частности, говорится, что планируемый в проекте анализ триад (отец, мать, ребенок) сокращает объем полезной геномной информации на одну треть, и поэтому вместо 1000 российских геномов фактически будет изучено 666 геномов.
|
О.П. Балановский отвечает А.А. Клесову на его рецензию статьи о генофонде балтов и славян. Тезисы А.А. Клесова о «подгонке генетических данных под лингвистику» и об отсутствии новизны оказываются взятыми с потолка. Примечательно, что критик выдает за выводы статьи то, что выводами совсем не является, и в то же время не замечает настоящих выводов. Очевидно, поверхностное знакомство со статьей, которую он берется рецензировать, рассчитано на таких же поверхностных читателей.
|
Древняя ДНК с Иберийского полуострова, показала, что генетически баски оказались потомками ранних европейских земледельцев и отчасти - местных охотников-собирателей. Представление об их длительной генетической изоляции подтвердилось.
|
Впервые генетикам удалось изучить древнюю митохондриальную ДНК Балканского полуострова – с территории Румынии. Это навело их на мысль о второй волне неолитической миграции в Центральную Европу через Балканы. Именно она внесла вклад в генофонд современных европейцев.
|
Йоганнес Мюллер – археолог, профессор Кильского университета (Германия), известный специалист по неолиту Европы, мегалитам и радиоугеродным датировкам. Публикуем его статью о проблемах воссоздания общественных идентичностей в археологии и генетике в переводе проф. Л.С.Клейна.
|
Профессор Гётеборгского университета Кристиан Кристансен дал интервью соредактору нашего сайта профессору Л. С. Клейну, В беседе специалистов подвергаются обсуждению некоторые заключения авторов статьи, вызывающие споры у археологов.
|
Эта наиболее полная работа по генофонду славянских и балтских народов подводит итоги многолетних исследований. Генетики и лингвисты проследили пути формирования генофонда всех групп славян и балтов одновременно по трем генетическим системам. Прослежено, какие местные популяции впитывал генофонд славян при их расселении по Европе: именно этот глубинный субстрат сформировал основные различия генофондов разных ветвей славян.
|
(краткий вариант)
Опубликована наиболее полная на сегодняшний день работа по изучению генофонда славян и балтов, в которой использован синтез генетики и лингвистики. При распространении по Европе славяне смешивались с местными популяциями, которые составили глубинный субстрат генофондов, отличающий разные ветви славян друг от друга.
|
Перевод статьи Кристиана Кристиансена, профессора университета Гётеборга в Швеции, ведущего специалиста по археологии бронзового века. В статье рассматриваются модели распространения индоевропейских языков в контексте социальных изменений, подтвержденных новыми археологическими данными.
|
Существуют различные точки зрения на прародину сино-кавказской языковой макросемьи (и включенных в нее дене-кавказских языков). Автор, развивая предложенную им несколько лет назад гипотезу локализации прародины дене-кавказской языковой общности в Восточной Евразии, предпринимает попытку показать, что и данные геногеографии приводят нас к такому же выводу.
|
В постсоветскую эпоху специалисты встретились с явлением, которое получило название «альтернативной истории». Что это за явление, чем оно вызвано, какими идеями оно питается и чему служит? Как специалистам следует на него реагировать? Об этом рассуждает доктор исторических наук В.А.Шнирельман.
|
Две статьи, вышедшие почти одновременно в Nature и Science, посвящены генетической реконструкции заселения Америки методами анализа полных геномов. Их выводы схожи. В статье команды Давида Райха (Nature), помимо основной миграции из Сибири, давшей начало всем коренным популяциям Америки, обнаружен – пока загадочный - «австрало-меланезийский след» у некоторых популяций южноамериканских индейцев. В статье команды Эске Виллерслева (Science) обнаружен тот же след, хотя его источник мог включать, кроме Австрало-Меланезии, еще и Восточную Азию.
|
Исследователи математически доказывают связь между лингвистическим и генетическим разнообразием в популяциях Европы. По их мнению, для изученных народов язык точнее, чем география, указывает на генетическое сходство популяций.
|
Группа исследователей из Калифорнии, применив передовые математические методы, получила для распада праиндоевропейского языка дату 6500–5500 лет назад, что соответствует гипотезе, согласно которой прародина индоевропейцев была в степи. Однако лексический материал, взятый ими для анализа, не выдерживает критики, поэтому достоверность результата в целом оказывается сомнительной.
|
В этой статье автор, профессор Л. С. Клейн, рассматривает ряд книг и статей по этногенезу, явно дилетантских, даже если их авторы и принадлежат к сословию ученых (обычно в науках, далеких от темы исследований). Украинские авторы упирают на украинское происхождение индоевропейцев, российские – на исключительную древность праславян и их тождественность с ариями.
|
Впервые по анализу древней ДНК удалось изучить, по каким генам и в каком направлении в популяциях Европы в последние 8 тысяч лет действовал естественный отбор. Под отбором находились аллели толерантности к лактозе, пигментации кожи и глаз, метаболизма, а также роста и веса.
|
Существует ряд методов обнаружения в геноме современного человека фрагментов ДНК, заимствованных из древних популяций. Среди них есть генетические варианты, имеющие приспособительное значение в изменившихся условиях внешней среды и оказавшиеся под положительным отбором.
|
В 2015 году вышла книга украинского профессора и членкора Украинской академии наук А. Г. Химченко с сенсационными выводами о прародине индоевропейцев. В рецензии на эту книгу профессор Л. С. Клейн оценивает ее как низкопробную халтуру, невысоко ставит и самого автора.
|
В геноме современного человека на территории Европы возрастом 37-42 тыс. лет найдено 6-9% неандертальской ДНК. Она была приобретена всего 4-6 поколений назад. Это означает, что метисация сапиенсов и неандертальцев случалась не только на Ближнем Востоке но и в Европе.
|
Критический анализ концепции происхождения индоевропейцев Т.В.Гамкрелидзе и В.В.Иванова предлагает историк Сергей Конча, научный сотрудник Киевского университета им. Шевченко.
|
Генетики секвенировали 102 древних генома и обнаружили динамичную картину перемещений, смешений и замещений популяций Евразии в бронзовом веке. По мнению авторов это дает ключ к загадке распространения индоевропейских языков.
|
Генетики показали родство «Кенневикского человека» с популяциями американских индейцев, а не с полинезийцами и айнами, как первоначально решили антропологи.
|
Анализ полногеномных данных современной популяции Египта и других африканских популяций привел генетиков к выводу о преобладании северного пути (через Египет) при выходе Homo sapiens из Африки.
|
Исследование генофонда Индии по полногеномной аутосомной панели GenoChip указало на преобладание в нем юго-западноазиатского компонента. Также ученые выяснили, что генетический ландшафт Индии довольно точно совпадает с географическим и лингвистическим делением её населения.
|
Полное секвенирование Y-хросомомы в 17 европейских популяциях показало, что от 2,1 до 4,2 тысячи лет назад почти по всей Европе началась Y-хромосомная экспансия — резкое увеличение эффективного размера популяции по мужской линии.
|
Публикуем аналитический обзор дискуссии "Спор о прародине индоариев" от историка, востоковеда, специалиста по древним и современным коммуникациям В.А.Новоженова. В обзоре разбираются аргументы "за" и "против" автохтонной концепции происхождения индоариев и анализируются многочисленные артефакты, свидетельствующие о возникновении и развитии колесных транспортных средств.
|
Публикуем статью доктора истор. наук Ю.Е.Березкина о том, что изучение распространения фольклорных мотивов может стать источником данных о миграциях популяций.
|
Накопленные данные по частотам микросаттелитных гаплотипов Y-хромосомы позволили исследователям обнаружить 11 крупных родословных кластеров в Азии. Их основателей можно считать отцами-основателями современной азиатской популяции, наряду с Чингисханом (Тимучином) и Гиочангом.
|
Публикуем аналитический обзор доктора истор. наук Л.С.Клейна дискуссии о происхождении индоариев. В данном обзоре Л.С.Клейн представил все обсуждаемые гипотезы, их аргументы и контраргументы, приводимые участниками дискуссии.
|
Дискуссия, которая развернулась в формате комментариев к заметке на сайте «Полное секвенирование отдельной гаплогруппы измеряет мутации и выявляет миграции» http://генофонд.рф/?page_id=2536. Тема происхождения индоариев, которая лишь косвенно относится к предмету исследования генетиков, вызвала бурные дебаты между сторонниками разных гипотез.
|
Перепечатываем беседу профессора Е.В Балановской с главным редактором журнала "Панорама Евразии"(Уфа) А.Т. Бердиным. Чем занимается наука геногеография? И почему ей необходимо решительно отмежеваться от ненаучных джунглей ДНК-генеалогии А. Клесова? Чем чреваты попытки дилетантов писать "народную генетическую историю"? Какие субъективные и объективные факторы позволили допустить квази-науку в здание Президиума РАН на карачаево-балкарской конференции?
|
Скифы – один из немногих бесписьменных народов древности, от которых до нас дошли и самоназвание, и достаточно подробные и в целом заслуживающие доверия сведения иноязычных нарративных источников. Тем не менее происхождение скифов остается предметом споров.
|
Изучив 456 секвенированных Y-хромосом из популяций по всему миру, исследователи уточнили и дополнили Y-хромосомное филогенетическое дерево, определили скорость мутирования на Y-хромосоме и обнаружили резкое снижение эффективного размера популяции по Y-хромосоме в районе 10 тысяч лет назад.
|
Исследователи нашли, что в современных популяциях европейцев и азиатов циркулируют фрагменты ДНК, составляющие около 20% генома неандертальцев. У азиатов их оказалось больше, чем у европейцев. Некоторые неандертальские аллели в геноме Homo sapiens поддерживались положительным отбором.
|
На основе полного секвенирования Y-хромосомной гаплогруппы G1 российские и казахские генетики построили детальное филогенетические дерево, вычислили скорость мутирования и генетически обосновали генгеалогию казахского рода аргынов.
|
Публикуем сокращенный вариант ветви дискуссии о гаплогруппах, языках и этносах к статье «ДНК-демагогия Анатолия Клесова», опубликованной в газете «Троицкий вариант-Наука». Обсуждение актуальных вопросов, затронутых в дискуссии, представляет интерес не только для ее участников, но и для широкого круга специалистов.
|
Представляем фрагменты из презентации доктора физико-математических наук, академика РАН Евгения Борисовича Александрова, председателя Комиссии по борьбе с лженаукой РАН «Лженаука в XXI веке в России и мире».
|
Продолжаем публиковать фрагменты из статьи археолога, этнолога и антрополога, доктора исторических наук Виктора Александровича Шнирельмана «Излечима ли болезнь этноцентризма? Из опыта конструирования образов прошлого — ответ моим критикам».
|
Публикуем фрагменты из статьи археолога, этнолога и антрополога, доктора исторических наук Виктора Александровича Шнирельмана «Излечима ли болезнь этноцентризма? Из опыта конструирования образов прошлого — ответ моим критикам», опубликованной в журнале «Политическая концептология» в 2013 году.
|
Урарту, скифы, аланы... Статья Л.С.Клейна в "Троицком варианте" о том, как народы бывшего Советского союза борются за право считаться потомками тех или иных древних народов.
|
«Битва за аланство» вспыхнула с новой силой. Некий анонимный документ, появившийся в интернете под видом резолюции карачаево-балкарской конференции 2014 года, уже привлек внимание общественности. Специалисты разбирают этот документ с позиций науки.
|
Впервые проведен полноценный тест современных филогенетических методов на лексическом материале лезгинской языковой группы.
|
Представляем интервью о проблемах этногенеза, опубликованное на сайте Полит.ру, с доктором исторических наук, археологом и филологом профессором Львом Самуиловичем Клейном и доктором биологических наук, генетиком и антропологом профессором Еленой Владимировной Балановской.
|
Слайд-доклад О.П.Балановского на междисциплинарной конференции в Звенигороде посвящен изучению древней ДНК, современных генофондов, а также сотрудничеству генетиков и этнографов.
|
Экспедиции в Крым проводились на протяжении четырех лет (2010-2013 годы) дружным международным коллективом – украинских и российских генетиков при активной поддержке и участии Меджлиса крымскотатарского народа и многих представителей крымских татар. Цель этой работы - реконструировать все составные части генофонда крымских татар.
|
Генетики изучили рекордное число образцов древней ДНК европейцев и нашли признаки миграции в центральную Европу из причерноморских степей около 4,5 тысяч лет назад. После появления новых генетических данных споры о происхождении индоевропейцев разгораются с новой силой.
|
Слайд-доклад Е.В.Балановской на междисциплинарной конференции в Звенигороде выявляет разногласия между генетиками и этнологами и предлагает конкретные шаги для их преодоления.
|
Чем занимается каждая из этих областей - популяционная генетика и генетическая генеалогия? На этот вопрос отвечают по-разному. В первом диалоге мы попробуем выяснить, как мы видим наши сферы действия.
|
Чем занимается популяционная генетика и генетическая генеалогия? На тот же самый вопрос, что и в первом диалоге, отвечают два известных представителя этих областей - Олег Балановский и Вадим Веренич.
|
Перепечатываем коллективную статью ученых в газете «Троицкий вариант-наука», обеспокоенных снижением иммунитета научного сообщества, допустившего дилетантское выступление А.Клесова на академическую трибуну.
|
В связи с выходом нового исторического журнала «Исторический формат», (о чем сообщил сайт Переформат .ру) мы обратились к историку О.Л.Губареву с просьбой прорецензировать те статьи этого журнала, которые близки его профилю.
|
|

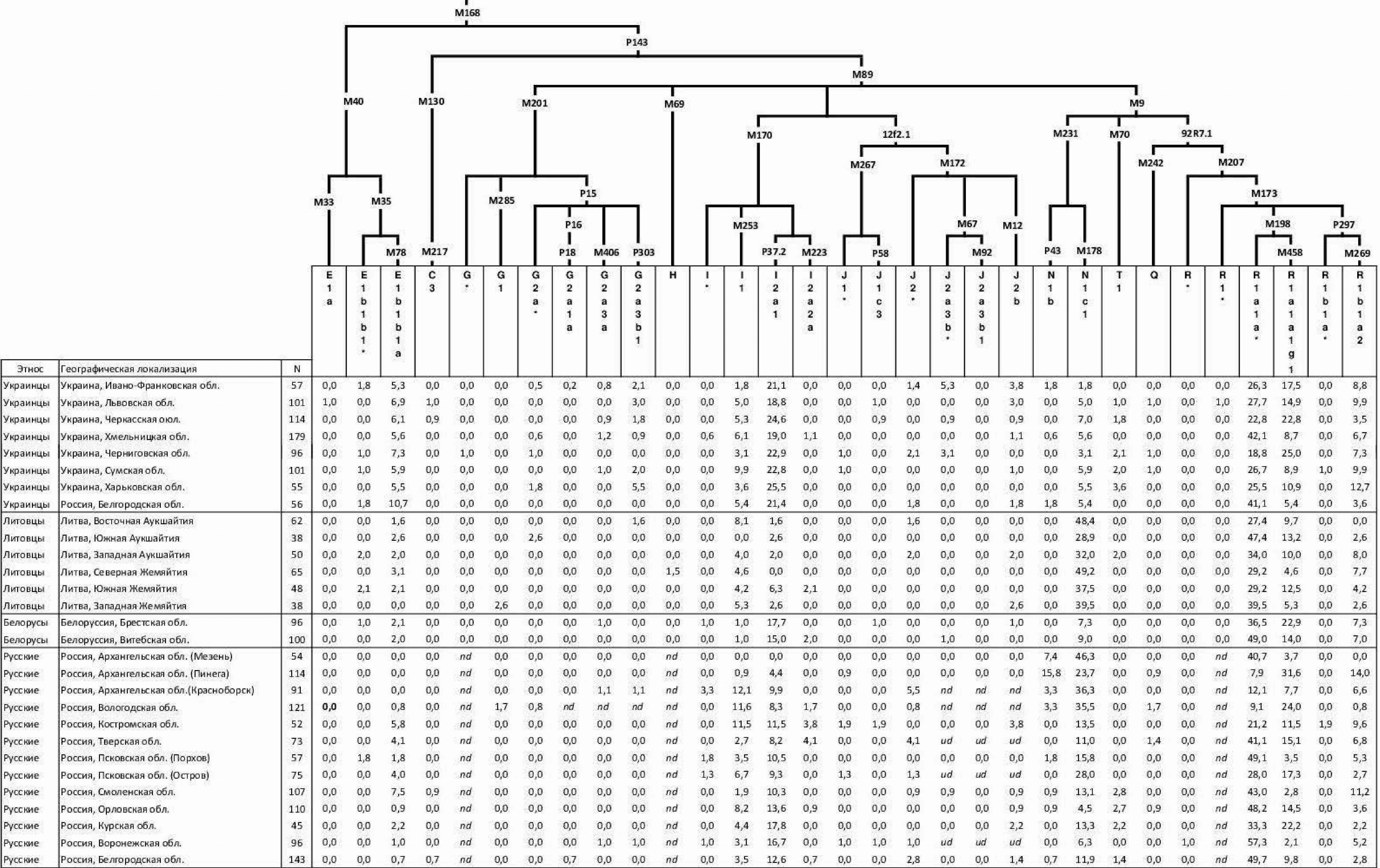
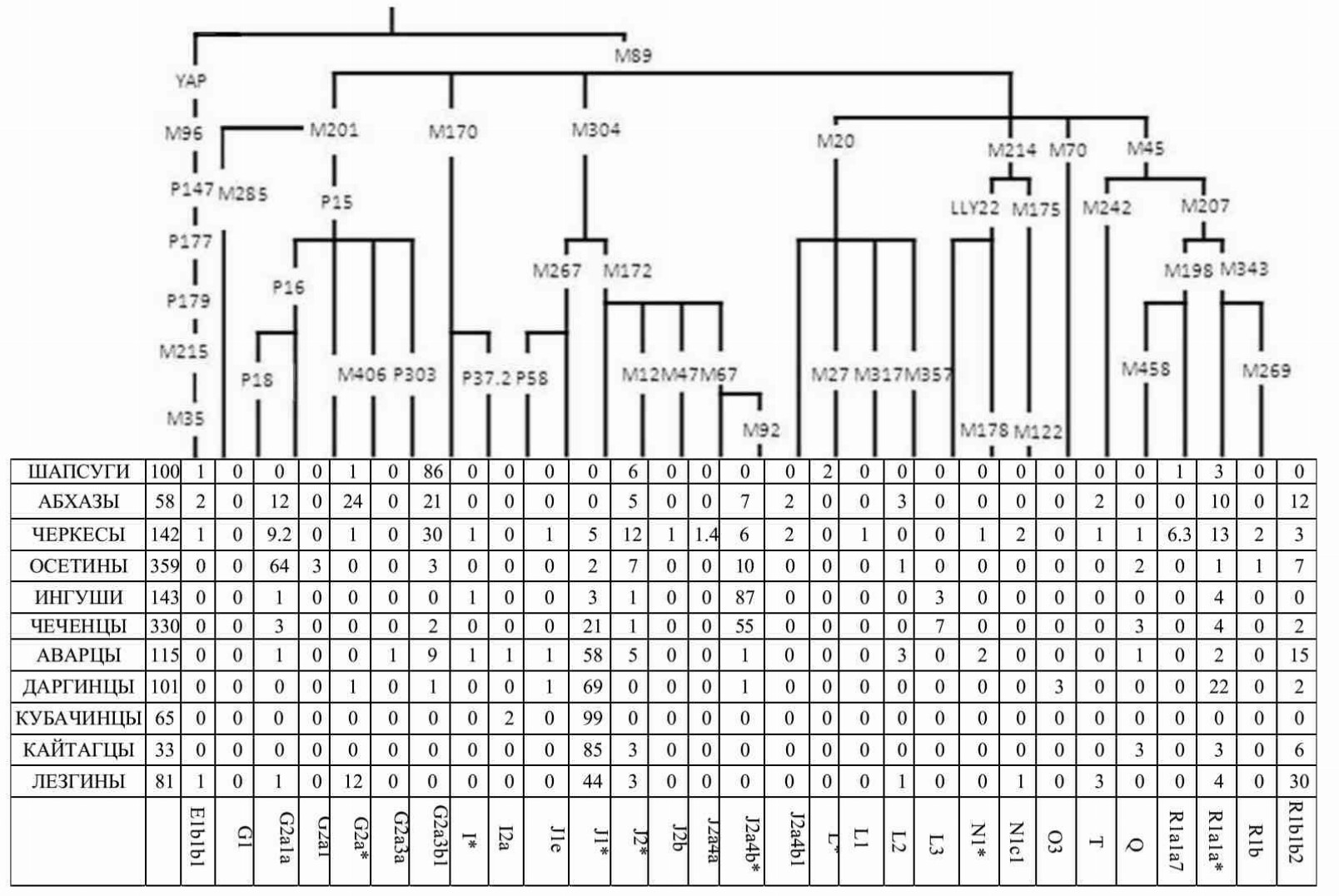

 Y-хромосомы в Европе.")
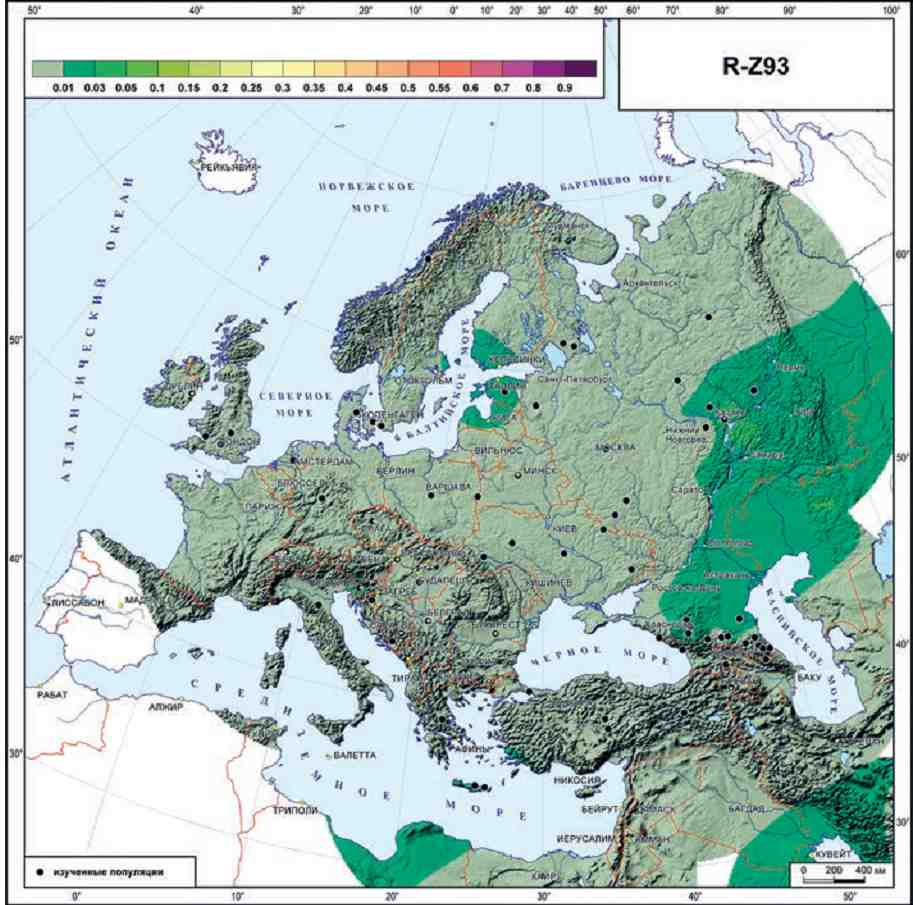




 Y-хромосомы в Европе.")









 Y-хромосомы в Европе.")



 Y-хромосомы в Европе.")
 Y-хромосомы в Европе.")






 Y-хромосомы в Европе.")
 Y-хромосомы в Европе.")






Исправленная карта Z93.
Благодарю Александра Штрунова и Владимира Таганкина, благодаря которым я обнаружил у себя эту ошибку в исходных данных (предыдущий вариант карты на самом деле относился не к Z93, а к Z93*).
Уважаемый Олег!
С интересом читаю Ваши статьи.
Был бы Вам очень благодарен, если бы Вы высказали своё мнение о «происхождении» гаплогруппы R1a1 у большинства современных тюркских народов (не считая северных тюрков-саха (якуты).
Ниже привожу мой вариант ответа на данную проблему.
Генофонд тюркских народов.
Российский генетик О.А. Балаганская в статье «ПОЛИМОРФИЗМ Y ХРОМОСОМЫ У ТЮРКОЯЗЫЧНОГО НАСЕЛЕНИЯ АЛТАЯ, САЯН, ТЯНЬ-ШАНЯ И ПАМИРА В КОНТЕКСТЕ ВЗАИМОДЕЙСТВИЯ ГЕНОФОНДОВ ЗАПАДНОЙ И ВОСТОЧНОЙ ЕВРАЗИИ» пишет: «Гаплогруппа R1a1a встречаясь во всех изученных популяциях с частотой от 4% до 61%, возможно, маркирует палеоевропеоидный субстрат, преобладавший на этой территории в скифо-сарматский и предшествующие периоды. Вариант R1b1b2(М269) напротив, встречается в изученных популяциях крайне редко, не достигая в среднем 1% (уровня полиморфизма).
ВЫВОДЫ:
— По маркерам Y хромосомы в генофонде тюркоязычных популяций Алтая, Саян, Тянь-Шаня и Памира (общий объем выборки N=1101 человек) выявлены три главных направления потока генов: западно-евразийское (маркируемое гаплогруппой R1a1a), восточно-евразийское (гаплогруппы C, D, O) и северо-евразийское (N1b, N1c1 и Q).
Миграции населения из Восточной Европы (гаплогруппы I и R1b1b2), Передней Азии (G и J) и Южной Азии (L) незначительны.
— Основная тенденция в формировании генофондов изученных тюркоязычных популяций выражается в замещении западно-евразийского (палеоевропеоидного) компонента восточно-евразийским (монголоидным) компонентом. Монголоидное влияние более выражено в степных и пограничных со степью районах, чем в горах, которые сыграли роль «барьера», и в генофонде которых сохранился вклад протоуральских и палеоазиатских племен (гаплогруппы N1b, Q).
— В генетическом пространстве изученные тюркоязычные популяции объединились в три кластера: Алтае-Саянский, степной и переходный между ними Памиро-Тяньшанский. По обеим системам маркеров Y хромосомы (SNP и STR) зафиксированы этногенетические связи киргизов с хакасами; шорцы наиболее близки к западно-евразийскому (европеоидному) кластеру; казахи
— Для наиболее частых гаплогрупп (N1b, N1c1, R1a1a, R1b1b1, Q, D) выявлены кластеры STR-гаплотипов, генетические датировки которых указывают на относительно молодой возраст генофондов народов Южной Сибири и Средней Азии (в среднем около 4000 лет)».
Этим самым О.А. Балаганская подтверждает появление первых тюрков в Сибири-4000 лет тому назад.
Российские генетики Е.В. Балановская , О.А. Балаганская , Л.Д. Дамба , Х.Д. Дибирова , А.Т. Агджоян ,Ю.В. Богунов , М.К. Жабагин , Ж.Т. Исакова , М.Б. Лавряшина , О.П. Балановский в статье «ВЛИЯНИЕ ПРИРОДНОЙ СРЕДЫ НА ФОРМИРОВАНИЕ ГЕНОФОНДА ТЮРКОЯЗЫЧНОГО НАСЕЛЕНИЯ ГОР И СТЕПНЫХ ПРЕДГОРИЙ АЛТАЕ-САЯН, ТЯНЬ-ШАНЯ И ПАМИРА» подводят итоги многолетних генетических исследований современных восточных тюркских народов.
Основные выводы российских генетиков:
-По маркерам Y-хромосомы в генофонде тюркоязычных популяций Алтае-Саян, Тянь-Шаня и Памира выявлены три главных направления потока генов: западно-евразийское (маркируемое гаплогруппой R1а1а);
Здесь укажем, что гаплогруппа R1а1а является доминирующей (средняя доля 45 %) у всех исследуемых современных тюркских народов (кроме якутов). R1а1а составляет у шорцев 60%,у тубаларов и алтай-кижи -50%. У остальных исследуемых тюркских народов колеблется от 24 до 57%. Вариант R1b1b2 встречается крайне редко, не достигая в среднем и 1%.
-центрально азиатское (C,D,O);
Необходимо отметить, что эти гаплогруппы отсутствуют у алтайцев хакасов и шорцев. В горах Алтая и Саян гаплогруппа С практически не встречается, за исключением самых южных территорий Алтая. У народов Южной Сибири по литературным данным (тувинцы, алтай-кижи, телеутам) частота С не более составляет 9%. О.А,Балаганская считает, что её распространение связано с широкой экспансией монгольских племен по степной полосе к югу от Алтайских гор. Гаплогруппа О и её субвариант О3, являющиеся основным компонентом генофонда в Восточной Азии (62%), встречены среди тюркских народов только на самом юге Алтая, в степных районах у казахов и с небольшим частотами у киргизов.
-северо-евразийское (N1b, N1c1, Q);
О.А.Балаганская считает, что «Об участии протоуральского и палеоазиатского населения в этногенезе населения Алтая можно судить по частоте распространения гаплогруппы Q, наиболее частой в Западной Сибири (в среднем около 32%). В генофонде алтайцев и хакасов гаплогруппа Q встречена нами с высокой частотой 17-18%. Также гаплогруппы N1c1 и N1(xN1c1), составляющие суммарно от 2 до 89 % в изученных группах, могут маркировать именно сибирскую составляющую в генофонде народов Алтае-Саян и некоторых среднеазиатских народов».
По мнению этих российских генетиков «основная тенденция в формировании генофондов тюркоязычных популяций состояла в постоянном замещении западно-евразийского субстрата (R1а1а-Гумбатов Гахраман) центральноазиатским (монголоидным) компонентом».
В 2012 году у азербайджанцев Ирана генетики обнаружили реликтовую гаплогруппу R1a (SRY10831.2), а также более древнюю предковую гаплогруппу R1a (M420) и (V.Grugni et al, 2012; Underhill et al,2014).
Американский генетик П.Андерхилл в статье, написанной в соавторстве с 32 ведущими генетиками мира, пишет: «Наши филогеографического данные приводят нас к выводу, что начальные эпизоды диверсификации R1a-M420 произошли в непосредственной близости от Ирана и Восточной Турции (Провинция Западный Азербайджан, Иран- Г.Г.) и мы оцениваем, что эта диверсификация произошла ~ 5800 лет назад. Из 24 R1a-M420 * (xSRY10831.2) хромосом в нашем наборе данных, 18 были отобраны в Иране ((Провинция Западный Азербайджан, Иран- Г.Г.), 3 были из восточной Турции. Аналогично, пять из шести наблюдаемых R1a1-SRY10831.2 * (xM417 / Page7) хромосомы были также из Ирана.
Благодаря распространенности базальных линий и высоким уровнем гаплогруппы разнообразия в регионе, мы находим убедительные аргументы в пользу Ближнего Востока (место возникновения R1a-Г.Г.), возможно, около современного Ирана, как географическое происхождение R1a». (Underhill et al,2014).
Исходя из новых генетических исследований ((Grugni et al (2012), Underhill et al (2014)) мы можем предположить, что примерно 6000 — 5500 лет назад часть древних тюрков носителей гаплогруппы R1a (M420)>(M198), (M417) > (SRY10831.2) в поисках новых пастбищ с Южного Кавказа передвинулись на Евразийскую равнину- R1a (SRY10831.2) > (Z93). Другая часть древних тюрков (предки современных азербайджанцев) осталась жить на Южном Кавказе. В дальнейшем, в результате повторной миграции, отдельные тюркские народы носители субкладов (Z93), (Z2125), (Z2123) возвратились на Южный Кавказ на территорию исторической прародины (киммерийцы, скифы, сарматы, аланы, гунны, кипчаки, огузы-сельджуки и др.) и в результате они вместе с аборигенными тюрками образовали современный азербайджанский народ.
Как известно гаплогруппа R1a является маркером распространения тюркского этноса и курганной культуры. Высокие частоты гаплогруппы R1a типичны для всех тюркских народов и их исторических соседей по Евразии, с которыми подвижные тюркские племена контактируют более 5000 лет.
Таким образом, можно предположить, что:
— большинство современных мужчин, носителей гаплогруппы R1a, являются потомками протюрков — выходцев из Южного Кавказа.
— генетические корни всех современных восточных тюркских народов находятся на Южном Кавказе. На территориях Восточнее Урала предки современных тюркских поселились 4000 лет тому назад.
Гахраман,
прежде чем начать обсуждение происхождения R1a у тюрков Сибири, надо обсудить принципиальные вопросы методологии. Потому что Ваше рассуждение: «Балаганская с соавторами встретили субварианты R1a у тюрок, Андерхилл с соавторами написал, что R1a в целом происходит из Передней Азии, Южный Кавказ относится к Передней Азии, азербайджанцы живут на Южном Кавказе, следовательно, все тюрки произошли от предков азербайджанцев», к сожалению, состоит из вопиющих логических ошибок. Думаю, они и Вам видны.
Например, из этой «логики» следует, что прототюрки — это эфиопы. Ведь R1a — субвариант гаплогруппы A’B, а ее происхождение тот же Андерхилл помещает на Эфиопское нагорье.
Также Вы напрасно приписываете Балаганской с соавторами вывод, что тюрки жили в Сибири 4000 лет назад. Из приведенной Вами цитаты видно, что эта датировка относится к кластерам гаплотипов, обнаруженных в популяциях этого региона, а вовсе не к самим этим популяциям. Поясню, в чем тут логическая ошибка: у тюрок Алтая, как и у всех людей, есть группы крови АВ0, которые есть и у бактерий; поэтому решайте — или тюрки Алтая происходят от бактерий, или наличие в популяции гаплотипов определенного возрасто ровно ничего не говорит о возрасте самой популяции.
Более того, в приведенной Вами цитате высказывается происхождение что эти конкретные кластеры гаплотипов были унаследованы местным населением от скифов (а вовсе не от тюрков).
Резюмирую: я считаю вполне возможным,что R1a пришла из Передней Азии (хотя вряд ли с Южного Кавказа, скорее из других частей Передней Азии). Но считаю совершенно невозможным, чтобы переднеазиатские носители этой гаплогруппы в то время говорили на тюркских языках. То есть я соглашусь, что Вы можете их считать одними из предков (не по языку, а по биологии) тюрков Сибири; но если хотите оценить, какова примерно доля именно этих предков среди всех предков тюрок Сибири, Вам достаточно заглянуть в статью Yunusbaev et al., 2015 на график ADMIXTURE и посмотреть доли «ближневосточного» (переднеазиатского, юго-западноазиатского) компонента. Я точно не помню, что предполагаю, что его 1-5%.
Гахраман, посмотрите: я потратил время, чтобы объяснить, что мне представляется неверным в Вашей логике, а Вы даже не снизошли до ответа. Вам это кажется нормальным?
Попробуем продолжить нашу «дискуссию».
Как известно, европеоидная гаплогруппа R1a1 выявлена также почти у всех современных тюркских народов:
Котоны (монголязычные уйгуры) -82,0 %,
Киргизы- 63,0 %,
Шорцы- 58,8%
Алтайцы- 53,0%
Татары- 34,1 %
Чуваши- 31,6%
Узбеки- 30,0%
Уйгуры- 28,6%
Хакасы 28,3 %
Карачаевцы — 27.54%
Башкиры- 26,3 %
Балкарцы — 25.74%
Азербайджанцы – 19,0%
Каракалпаки- 18,2%
Тувинцы 14, 0%
Кумыки- 13,2%
Гагаузы — 12,5%
Турки- 6,9%
Туркмены — 6,7 %
Казахи — 4,0%
Якуты — 3,2%
Кроме того генетики выявили, что «восточноазиатские» гаплогруппы (N, O, C, D), присущие монголидным народам, у «западных» тюркских народов (азербайджанцев, турков, туркмен, гагаузов, карачаевцев, балкарцев, кумыков) не обнаружены.
Ознакомившись с этими данными по ДНК генеалогии мы можем предположить, что гаплогруппа R1a1 изначально была древнетюркской гаплогруппой. А тот факт, что у всех мужчин, захороненных в евразийских курганах, обнаружена эта гаплогруппа, является ещё одним доказательством того, что так называемые андроновцы, афанасьевцы, тагарцы, «пазырыкцы», скифы, сарматы, аланы были тюркскими народами и являются древними предками современных тюрков. Об этом в своих исследованиях пишут большинство зарубежных и российских генетиков. Так, например, российские исследователи Волков В.Г, Харьков В.Н., Штыгашева О.В., Степанов В.А в статье «Генетическое исследование хакасских и телеутских сеоков» пишут: «Практически все гаплотипы R1a1 у хакасов и шорцев входят в единую группу и отличаются от гаплотипов R1a1 других этносов, но имеют явное сходство с гаплотипами носителей тагарской археологической культуры. Таким образом, данные сеоки являются прямыми потомками тагарцев».
А в другом исследовании по генетике населения Южной Сибири «Эволюция и филогеография линий Y-хромосомы человека» генетики В.А. Степанов, В.Н. Харьков, В.П. Пузырев пишут, что «Большую часть в спектре вариантов Y-хромосомы в Южной Сибири занимает R1a1 (от 12 % у тувинцев до 55 % у южных алтайцев), носители которой – вероятно, древнеевропеоидное население этого региона – проникли сюда с миграциями по степной зоне Северной Евразии в эпоху от раннего неолита до бронзового века».
Археологи выявили комплекс взаимосвязанных и относительно мобильных культур,
проживающих в древности на территории евразийских степей (курганная культура).
Как известно у монголов доминирует гаплогруппа С (M217), у тунгусов -N1a (M128),
у корейцев- D4, у японцев-D2 , китайцев-O (M175).
Антрополог Т. А. Чикишева в заключении диссертации «Динамика антропологической дифференциации населения юга Западной Сибири в эпохи неолита — раннего железного века» пишет: «Южная евразийская антропологическая формация явилась антропологическим субстратом всех известных к настоящему времени в Алтае-Саянском регионе автохтонных культур. Она вошла в состав некоторых популяций афанасьевской культуры… Очевидные миграционные импульсы на территорию Горного Алтая с юга из среды скотоводческого населения Передней или Средней Азии прослеживаются по антропологическим данным, начиная со II тыс. до н.э., и усиливаются в эпоху ранних кочевников… В антропологическом составе носителей пазырыкской культуры Горного Алтая выявляется европеоидный компонент, генетически восходящий к скотоводческому населению северных районов Передней Азии и южных районов Средней Азии».
Мы можем предположить, что тюрки скотоводы с конца неолита и в железном веке принесли на просторы Евразии неизвестную ранее культуру, которая имела ряд специфических черт, такие как курганные захоронения, одомашненные животные (овца, коза, кр. рогатый скот, лошадь, верблюд) и металлургию. Почти все образцы из бронзового и железного века погребенных в курганных захоронениях Южного Кавказа, а также в причерноморских и прикаспийских погребениях (ямная, катокомбная, андроновская), вплоть до погребений афанасьевцев в Красноярском районе в южной части Сибири и таримцев на территории современной Уйгурии, принадлежало населению с общей гаплогруппой R1a1-M17. Что также подтверждает миграции европеоидных древних тюрков с территории Южного Кавказа на восток.
Гахраман,
что Вы понимаете под термином «тюрки»? Надеюсь, что Вы следуете общепринятому пониманию (народы, говорящие на тюркских языках). Если же что-то другое, то поясните. А то у Вас получаются анахронизмы наподобие влияния тюрок на шумеров (неандертальцев, австралопитеков, динозавров и далее в глубь времен).
Чтобы дискуссия получилась, надо отвечать на высказывания собеседника. Я на Ваши отвечаю, а Вы мои игнорируете и продолжаете говорить свое. В качестве последнего аванса последний раз отвечу на Ваши соображения, и жду того же от Вас.
1) Гипотезу о Ra1 как заметной части генофонда прототюрков выдвинуть можно. Но наличие R1a у таких народов, как турки, азербайджанцы, гагаузы, карачаевцы, балкарцы и т.д., (у которых тюркские языки являются недавним приобретением, почти не отразившемся на генофонде, что видно из антропологии и аутосомного генома) свидельствует против такой гипотезы.
2) Наличие R1a в степях до того времени, как тюркские языки вообще возникли (а это время хорошо датируется глоттохронологией) вообще не имеет отношения к истории тюрков, хотя и указывает, что современные степняки (тюрки) вобрали в себя часть генофонда прежних степняков (скифов), безотносительно тому, на каком языке эти самые скифы говорили.
3) Миграции из Передней или Средней Азии в степи Сибири, о которых пишут цитируемые Вами антропологи, Вы связываете с тюрками без каких-либо оснований (по крайней мере Вы этих оснований не привели). В степи Сибири кто только не мигрировал (вплоть до русских)…
1) что именно и чему именно противоречит?
2) А из перечисленных Вами народов — предки украинцев и мордвы наверняка, русских — только южных, марийцев и литовцев — лишь в малой степени, — осетин — скорее всего, в очень малой степени.
3) Приведенные Вами авторы ни в коей мере не утверждали, что эти мигранты были тюрками.
Констатирую, что диалога не получилось, поскольку Вы не только не отвечаете, но даже не вчитываетесь в то, что отвечают Вам.
Что это за степняки R1a? Различные ветви Z93+?
Вот с этого надо было начинать. То есть Вы считаете, что эта популяция была предковой для тюркских народов, но согласны с тем, что эта популяция говорила вовсе не на тюрских языках. Или все же считате, что тюркские языки существовали 8 тысяч лет назад?
Предполагаемые потомки скифов.
Дополнительные аргументы.
Как известно, до сих пор, к сожаленью, нет специального исследования, посвящённого дописьменной истории древних тюрков.
Историческую прародину древних тюрков можно определить на основании сравнительного анализа современных и древних тюркских языков, путём сопоставления полученных результатов с данными археологии, антропологии, этнографии и исторических материалов.
Учёные утверждают, что каждая гипотеза может претендовать на правдоподобность только в том случае, если она имеет достаточное основание. Закон достаточного основания, выражает одну из общих черт правильного мышления — его обоснованность. Этот закон требует, чтобы наши суждения о предмете и его свойствах были не голословны, а базировались на достоверных аргументах
Известный российский лингвист Б.А.Серебренников писал по этому поводу следующее: «Всякая мысль, чтобы стать несомненной, должна быть обоснована другими мыслями, истинность которых доказана или самоочевидна»
Серебренников Б.А. Проблема достаточности основания в гипотезах, касающихся генетического родства языков // Теоретические основы классификации языков мира: Проблемы родства. М., 1982.
Так, как я, как наверно, и любой другой исследователь, одновременно не могу быть высококвалифицированным специалистом в таких разных разделах науки, какими являются лингвистика, археология, антропология, этнография, история, религия и др, то для обоснования своих мыслей, я постарался как можно максимально использовать исследования известных мировых специалистов по вышеназванным разделам науки.
Ответ Вадиму Созинову.
Несмотря на Ваш высокомерный, а местами и …,так скажем, оскорбительный ответ: («можно навыдергивать сотни цитат, …Ваше незнание такой научной дисциплины как антропология, … Ваше последующие додумывания за них», «мне кажется для вас это будет бесполезно!…», Бред!», попробую довести до Вашего сведения, видимо, неизвестные Вам высказывания известных ученых по рассматриваемой нами теме:
В.П. Алексеев: «Из факта древности и автохтонного формирования кавкасионского типа уже был сделан вывод, что осетины представляют собой потомков местных этнических групп, населявших Центральный Кавказ с глубокой древности».
В.П. Алексеев. Историческая антропология и этногенез». 1989.
Г.Ф.Дебец: «В древнейших погребениях Самтаврского и Мингечаурского могильников находят резко выраженные длинноголовые узколицые европеоидные черепа, по типу сходные больше всего с представителями современных длинноголовых вариантов каспийского типа (к данному типу относятся тюркские народы-азербайджанцы и кумыки- Гумбатов Гахраман).
Народы Кавказа, Том 1, Академии наук СССР, 1960.
А.А.Казарницкий: «Лептоморфный краниологический комплекс, доминировавший в азово-каспийских степях в течение средней и поздней бронзы, находит ближайшие морфологические аналогии среди краниологических серий Закавказья».
А.А.Казарницкий. «Население азово-каспийских степей в эпоху бронзы (антропологический очерк). СПб.: Наука. 2012.»..
О.Исмагулов: «Основная масса местных насельников тюркского периода Казахстана характеризовалась европеоидными чертами. Если сравнить формирование лингвистической и антропологической общностей, то население, прежде всего, стало говорить на тюркских языках и лишь значительно позднее стало монголоидным. Преобладание монголоидных элементов в разных пропорциях в физическом облике местных насельников наблюдается главным образом в монгольский период».
О.Исмагулов. «Этническая антропология Казахстана».1982.
К.Н. Солодовников, С.С. Тур: «Антропологический состав населения елунинской культуры неоднородный и формируется в результате взаимодействия двух компонентов: местного промежуточного европеоидно¬монголоидного и пришлого европеоидного «средиземноморского». Первый компонент представлен в женской группе, второй — преимущественно в мужской».
К.Н. Солодовников, С.С. Тур. «Материалы к кранеологии эпохи ранней бронзы Верхнего Приобья». 2003.
А Джеймс Патрик Мэллори: «Прослеживая распространение русского языка по территории бывшего СССР, допустимо предположить, что оно шло из европейской части России с ~ 1100 до 1900 г. н. э. Но было бы хронологически абсурдно связывать это с носителями ямной культуры (~ 3000 – 1800 гг. до н. э.) и других, еще более восточных культур, например, афанасьевской на Енисее/Алтае».
Дж. П. Мэллори. «Индоевропейский феномен». 1999.
А.Н.Бернштам: «Прослеживая по археологическим памятникам историю развития кочевых обществ и выявляя автохтонный процесс их развития, мы приходим к выводу, что там, где начиная с эпохи бронзы, шел процесс формирования кочевого общества, там конечным результатом процесса являлся тюркский этногенез».
А.Н. Бернштам. Проблемы истории Восточного Туркестана. ВДИ. 1947. № 2.
Д.Г.Савинов: «В памятниках пазырыкской культуры Горного Алтая впервые появляется ряд элементов древнетюркского культурного комплекса. Это – сопроводительные захоронения коней, расположенные в могильной яме на приступке с северной стороны, и вертикально вкопанные камни с восточной стороны некоторых больших и малых пазырыкских курганов – прообразы будущих камней-балбалов…В настоящее время таких материалов уже «накоплено» вполне достаточно для того, чтобы поставить вопрос о выделении «скифского» и «хуннского» пластов (или компонентов) в древнетюркском культурогенезе. Целый ряд предметов материальной культуры и искусства, зародившись в эпоху ранних кочевников, продолжали существовать без значительных изменений и позже, органически «вписавшись» в культуру эпохи раннего средневековья… Долгое время, главным образом, благодаря неоднократному упоминанию в письменных источниках, установка камней-балбалов считалась одним из наиболее характерных элементов древнетюркского погребально-поминального комплекса. В настоящее время установлено, что камни-балбалы как вид культовых памятников появляются ещё в конце эпохи бронзы. Ряды камней-балбалов, отходящие на восток, установлены у больших и малых курганов пазырыкской культуры на Алтае». (150)
Д. Г. Савинов. О «скифском» и «хуннском» пластах в формировании древнетюркского культурного комплекса. 1998. С. 130-141.
Дмитриева Л. В.: «Получившие общетюркские обозначения растения и их части, вероятно, входили в ту ботаническую среду, которая окружала древних тюрков на их прародине. Они жили в районах с преобладанием деревьев (а именно березы, яблони), злаковых и трав, диких-гороха и лука, где могли произрастать просо, пшеница, ячмень».
Дмитриева Л.В.: «Лексика, относящаяся к животному миру, является одной из древнейших групп в тюркских языках. Названия домашних животных оформились в очень раннюю эпоху, в эпоху совместного проживания предков современных тюркских народов, то есть задолго до начала н.э.».
Дмитриева Л.В. «Из этимологий названий растений в тюркских, монгольских и тунгусо-маньчжурских языках». 1971. Стр.187.
Дж. Клоусон: «Результаты применения лексикостатистических методов к оценке «алтайской» теории можно суммировать следующим образом: После исключения слов, которые наверняка можно признать заимствованными, общие элементы в тюркском и монгольском основном словаре составят не более 2% от основного словаря, причем эти общие слова легче объяснить как заимствования, чем как свидетельство генетических связей…Тюркские языки и маньчжурский, по всей очевидности, не связаны генетически, так как их основной словарь не совпадает….Общие элементы в монгольском и маньчжурском основном словаре не превысят 3,5% от всего лексического состава, причем эти слова могут быть легче объяснены как заимствования, чем как свидетельство генетических связей… Следовательно, «алтайская» теория неправомерна».
Дж. Клоусон. «Лексикостатистическая оценка алтайской теории». Вопросы языкознания», 1969, № 5.
Марио Алинеи: «В четвертом тысячелетии до нашей эры курганные народы распространились по всей области к северу от Черного моря, по северной Европе, и вероятно к востоку от естественного барьера Уральских гор. Слово курган на тюркском языке означает холм или могила. Курганная культура характеризована ямными могилами или курганами, т.е. специфическим методом похорон. Самые ранние курганы находятся к северу от Черного моря, откуда они распространяются примерно к 2000 г. до н.э по Центральной Европе, пересекая Днепр… Как известно слово курган не русского, не славянского, и не индоевропейского происхождения, а заимствование из тюркских языков… Ямная культура, согласно моей гипотезе, является тюркскими, и это означает, что тюркские народы были первыми, кто успешно приручил лошадей, и передал это новшество соседним народам».
М.Алинеи. Курганная культура. 2003.
В.А. Степанов, В.Н. Харьков, В.П. Пузырев: «Большую часть в спектре вариантов Y-хромосомы в Южной Сибири занимает R1a1 (от 12 % у тувинцев до 55 % у южных алтайцев), носители которой – вероятно, древнеевропеоидное население этого региона – проникли сюда с миграциями по степной зоне Северной Евразии в эпоху от раннего неолита до бронзового века».
В.А. Степанов, В.Н. Харьков, В.П. Пузырев. «Эволюция и филогеография линий Y-хромосомы человека». 2006.
Волков В.Г, Харьков В.Н., Штыгашева О.В., Степанов В.А: «Практически все гаплотипы R1a1 у хакасов и шорцев входят в единую группу и отличаются от гаплотипов R1a1 других этносов, но имеют явное сходство с гаплотипами носителей тагарской археологической культуры. Таким образом, данные сеоки являются прямыми потомками тагарцев».
Волков В.Г, Харьков В.Н., Штыгашева О.В., Степанов В.А. «Генетическое исследование хакасских и телеутских сеоков». 2010.
Надеюсь, уважаемая редакция популярного сайта «Генофонд Р.Ф.» не поручит господину Созинову отвечать на данный мой пост. Заранее благодарю.
Уважаемая редакция никому ничего не поручает — на сайте для всех свобода высказывания.
Но не могу не согласиться с В. Созиновым, что Ваша манера цитирования отражает Вашу сверх-ангажированность. Приводимые Вами цитаты говорят совсем о другом, чем Вам хотелось бы.
Никакая редакция мне сказать это не поручала:).
Елена Балановская:
«Но не могу не согласиться с В. Созиновым, что Ваша манера цитирования отражает Вашу сверх-ангажированность».
ангажированный —поступающий каким либо образом в результате того, что был вовлечён кем либо в какое либо дело, привлечён к активному участию в чём либо, подкуплен предоставленными услугами, льготами и т.п.; необъективный.
http://dic.academic.ru/dic.nsf/ushakov/739362
Опять вместо диалога-ярлык.
Елена Балановская:
«Приводимые Вами цитаты говорят совсем о другом, чем Вам хотелось бы».
У Вас как я вижу, какой-то особый дар. Вы, судя по Вашим словам, даже можете влезть в мой мозг и увидеть там, какие-то затаенные мысли. Но в отличие от многих, я всегда говорю, то, что хочу сказать, а в научных спорах опираюсь на высказывания специалистов, которых, к сожалению, в последнее время очень мало.
Тюркские народы (сегодня вторые по численности в России), а Вы, к сожалению, ничего не знаете об этой этнической общности, которая и сегодня живет там, где жили их предки начиная с IIIтыс. до н.э. (евразийская равнина). На этой обширной территории от Дуная до Алтая в курганах лежат их далёкие предки. У которых тюркская гаплогруппа R 1a.
«АНГАЖИРОВАННОСТЬ — от французского «вовлеченность». Быть ангажированным — значит быть вовлеченным. Принять сторону… По Сартру, ангажированность — это сознательно выбранная общественная позиция». (с)
Общественная. Потому это не ярлык, а противопоказания для научных изысканий.
Мысли не затаенные, а ярко выражают Вашу «сознательно выбранную общественную позицию».
«…а в научных спорах опираюсь на высказывания специалистов, которых, к сожалению, в последнее время очень мало».
Правду сказали — очень мало настоящих ученых, которые бы отражали Вашу «сознательно выбранную общественную позицию».
О генофонде тюркоязычных народов мы знаем немало — изучаем народы от Крыма через Кавказ, Малую Азию, Закавказье, Урал, Западную Сибирь (вот книжка вышла про сибирских татар только что, где есть и наша глава про их генофонд), Южную Сибирь и Восточную до Центральной Азии. И поэтому кое-что можем сказать (не зря Ваш фильм пестрит вырезками из наших статей — но отчего-то без ссылок на первоисточники).
Например, что тюркоязычные народы с точки зрения генетики не представляют собой «общность».
И что R1a — не тюркская гаплогруппа.
Нехорошо присваивать общее достояние.
Здравствуйте, Гахраман. Вы меня рассмешили, что кто-то, якобы, даёт мне какие-то поручения.
И опять двадцать пять, это я к тому, что снова много «известных для меня» цитат.
Ещё раз повторюсь, никто, из приведенных вами авторов, не связывает прародину, родину тюрков, тюркских языков с Азербайджаном, Передней Азией, и одним из вариантов средиземноморской расы. И дальнейшие ваши измышления, и есть додумывание.
Не наука, а идеология налицо! Одна из частей Передней Азии = родина изначальных, истинНых носителей тюркских языков = одна из ветвей средиземноморской расы = дальнейшая миграция на восток = смешение с монголоидами = возвращение домой на запад = и при этом на протяжение веков практически неизменяемый язык!?
При этом, настоящие учёные-исследователи пишут, и употребляют, в своих работах такие обороты речи, как: возможно, по имеющимся данным, предположительно, и т.д., но никогда не говорят и не пишут со 100% уверенностью.
Что и выдаёт в вас, идеолога!
Уважаемый Олег, посмотрите 5 минутный фильм по генетике тюркских народов (без шумеров) и может быть Вам моя точка зрения покажется более аргументированной.
https://www.youtube.com/watch?v=YB7KyV1jo2o
Заранее благодарю.
Ого. Такого я еще не видел. Вы правы, это не шумеры, это похлеще будет.
Обратите внимание, что название фильма «Генетика против алтайской теории», при этом даже не упоминается специально посвященная тюркам статья генетиков (Yunusbaev et al., 2015) в которой как раз миграции тюрков с алтае-монгольской прародины и продемонстрированы.
Олег Балановский сказал(а):
«Ого. Такого я еще не видел. Вы правы, это не шумеры, это похлеще будет».
Олег, «юмор» Ваш, мягко говоря, весьма своеобразный.
«Обратите внимание, что название фильма «Генетика против алтайской теории», при этом даже не упоминается специально посвященная тюркам статья генетиков (Yunusbaev et al., 2015) в которой как раз миграции тюрков с алтае-монгольской прародины и продемонстрированы».
Я ничего против Юнубсаева не имею, но в большинстве случаев, люди( писатели, исследователи. «творцы») на госоплате не могут писать. творить, создавать, против генеральной линии госнауки.
Вы имеете в виду, что создание этого фильма финансировалось госнаукой Азербайджана? Сочувствую ученым этой страны.
В России-то запрета противоречить официальной линии, к счастью, уже полвека как нет (после окончания лысенковщины). По крайней мере за генетику могу ручаться.
«Вы имеете в виду, что создание этого фильма финансировалось госнаукой Азербайджана? Сочувствую ученым этой страны.»
Олег, этот фильм смонтировал я. Как и остальные 35 фильмов. https://www.youtube.com/user/patriot2066. Если будет желание или время (например,при поездках на симпозиумы). У пенсионера с 40 летним стажем работы много свободного времени.
«В России-то запрета противоречить официальной линии, к счастью, уже полвека как нет (после окончания лысенковщины). По крайней мере за генетику могу ручаться».
Олег, я рад за российскую науку.
Учитывая Ваше отрицательное отношеник к большим текстам, направляю Вам сокращённый вариант своих аргументов.
Гордон Чайлд пишет: «В гатах Зороастра, где зарисовки сельской жизни представлены в изобилии, главным достоинством ария считается занятие земледелием, а кочевники проклинаются, как грабители из Турана… В VIII веке до н.э. народ, который ассирийцы назвали ашгузай, a греки — скифами, пересек Кавказ, чтобы вторгнуться в Месопотамию. Многие исследователи считают, что скифы были иранцами. Данные лингвистики, которые ограничиваются несколькими именами собственными, в основном более позднего времени, являются не очень убедительными. Однако археологам этот народ хорошо известен. Без всякого сомнения, скифское искусство испытало на себе сильное иранское влияние, но нельзя сказать, что оно сформировалось под влиянием искусства какого-нибудь определенного иранского народа, например персов, — оно имеет свою собственную уникальную индивидуальность. Решающее значение для этнической идентификации скифов приобретает изучение их погребального обряда. Он сильно отличается от того обряда, который был присущ иранцам или индийцам, как, впрочем, и любому другому индоевропейскому народу. Эти обычаи описаны Геродотом, их следы найдены при раскопках многих курганов, но они совершенно неарийские. Они находят точные соответствия у кочевников Монголии, которых никак нельзя отнести к ариям, причем они существуют у них на протяжении многих веков, как было показано Миннзом. Он считает скифов предшественниками гуннов, татар и печенегов, и это не вызывает сомнения».
С. Е. Малов: «За пять веков до н. э. тюрки жили там же, где они живут главным образом (с малыми исключениями) и теперь. В Европе древнейшими местами обитания тюрков были: р. Дунай, нижнее и среднее течение Волги, бассейн Урала.».
В. И. Рассадин: «Специалистами по этимологии тюркских языков убедительно доказано, что все видовые и половозрастные названия пяти видов скота (лошади, крупный рогатый скот, верблюды, козы и овцы – Г. Г.), традиционно разводимые тюрками и представленные как в древнетюркском языке, так и в современных, унаследованы языками — потомками от общетюркского праязыка вместе с самим степным скотоводством номадного типа, пришедшим из глубины тысячелетий».
И.А.Джавахишвили: «На основании измерений всех черепов, обнаруженных в древнейших захоронениях Кавказа, – было установлено, что ранее в нашей стране жили длинноголовые, т.е. долихокефалы. Профессор Вирхов (известный немецкий антрополог-Г.Г.) отмечал, что нынешние армяне и грузины не имеют ничего общего с древнейшим населением Кавказа. Следовательно, по сути, армяне и грузины не являются первыми местными поселенцами. Когда они пришли в эту страну, здесь уже жили потомки другого народа».
М.Г.Абдушелишвили: «В палеоантропологических материалах из Восточного Закавказья нет данных, свидетельствующих о появлении каких-либо иных форм, отличающихся от местных, наоборот, все свидетельствует о преемственности населения, проживающего с незапамятных времен на данной территории. Поэтому мы не находим в палеоантропологических материалах оснований для утверждения, что каспийский тип сформировался в процессе заселения Кавказа с юго-востока».
Г.Ф.Дебец: «В древнейших погребениях Самтаврского и Мингечаурского могильников (Южный Кавказ-Г.Г.) находят резко выраженные длинноголовые узколицые европеоидные черепа, по типу сходные больше всего с представителями современных длинноголовых вариантов каспийского типа».
В.В.Бунак: «В степных пространствах Средней Азии и Южной Сибири в глубокой древности сложилась особая группа типов, отличающаяся от средиземной группы более широким и высоким лицом, узким носом, массивным надбровьем, более прямыми волосами, -евразийская ветвь. К ней следует отнести древнейшие типы, известные по краниологическим материалам афанасьевской и андроновской культур, и, вероятно, некоторые другие, более западные. В современную эпоху евразийская группа представлена каспийским типом у туркмен и азербайджанцев … В древности евразийские варианты имели большое распространение и в составе скифских и сарматских племён продвинулись в Переднюю Азию, на Кавказ и Восточную Европу вплоть до Нижнего Дуная».
Тур С.С.: «Появление в данном регионе долихокранного европеоидного типа, именуемого также гиперморфным восточносредиземноморским, связывается с миграцией скотоводческих племен из Средней или Передней Азии. Современные популяции североалтайского антропологического типа, к которым относятся северные алтайцы, телеуты, горные шорцы, а также барабинские татары позднего времени, являются потомками носителей пазырыкской культуры. Характерный для них краниологический комплекс формируется на территории Горного Алтая в результате длительного смешения и нивелировки особенностей двух основных компонентов различного происхождения — брахикранного, с умеренно выраженными монголоидными особенностями и низким, по монголоидному масштабу, лицом и долихокранного европеоидного, с высоким и широким лицом. Первый из них встречается на территории Горного Алтая с эпохи энеолита, второй появляется в эпоху ранней бронзы в результате миграции скотоводческих племен предположительно из Средней или Передней Азии».
Олег, могли бы Вы высказаться об испанских тюрках-пасиего?
Генетическая характеристика пасиего.
Пасиего имеют своеобразную культуру, а также необычный для Испании генетический набор.
Среди пасиего распространены гаплогруппы R1b, R1a, G2 и E1b.
Гаплогруппы R1a и G2 не характерны для населения Испании и всей Западной Европе.
Как полагают генетики, носители гаплогрупп R1a и G2 пришли на территорию Западной Европы с Южного Кавказа во времена Мегалитической культуры (III тыс. до н.э.).
У пасиего около 80% их ДНК совпадает с восточноевропейскими тюркскими народами (азербайджанцы, кумыки, карачаевцы, башкиры и другие). По сравнению с другими народами Западной Европы пасиего имеют высокие частоты R1a (SRY10831.2)- 18,0%. Как известно,
у азербайджанцев Ирана генетики обнаружили аналогичную реликтовую гаплогруппу R1a (SRY10831.2), а также более древнюю предковую гаплогруппу R1a (M420) и (V.Grugni et al, 2012; Underhill et al,2014).
http://www.proza.ru/2014/09/16/410
https://www.youtube.com/watch?v=2YT2Nz_P3xA
Видимо, Олег потерял интерес к данной дискуссии. Что же, может быть на «Переформат.ру» можно будет, наконец-то, выяснить кого можно считать потомками евразийских скифов.http://pereformat.ru/2015/12/nasledniki-ariev/
Здравствуйте Гахраман.
Во-первых, то, что вы привели мнение археолога Гордона Чайлда — это всего лишь его мнение, а не всего научного мира.
Во-вторых, Малов С.Е. не приводит никаких лингвистических убедительных доказательств того, что тюркские языки были распространены с 5 в. до н.э. в области от Дуная до Урала.
В-третьих, сказанное Рассадиным В.И. вообще не понятно для чего здесь вставленно, так как все мы унаследовали от своих языковых предков понятия видов скота и т.п.. И что, тюрки здесь какое то исключение?
В-четвертых, выкладки цитат от антропологов, у которых эти антропологические типы никогда не связывались с предками тюрков, вообще бессмысленны, т.к. это лишь показывает Ваше незнание такой научной дисциплины как антропология, и Ваше последующие додумывания за них. Это называется свалить все в кучу. Таким же способом можно навыдергивать сотни цитат против ваших выкладок, но мне кажется для вас это будет бесполезно!
По поводу пасиего.
G2 может быть от неолитчиков, у R1а сделаны онп только верхнего уровня, то есть опять сплошные додумки, как и в случае с недотестированными М420 у азербайджанцев Ирана, в прошлом говоривших на одном из иранских языков!
И где общего 80 % генофонда между пасиего и каквказскими народами? Бред!
Елена Балановская:
18.02.2016 в 23:04
О генофонде тюркоязычных народов мы знаем немало — изучаем народы от Крыма через Кавказ, Малую Азию, Закавказье, Урал, Западную Сибирь (вот книжка вышла про сибирских татар только что, где есть и наша глава про их генофонд), Южную Сибирь и Восточную до Центральной Азии. И поэтому кое-что можем сказать (не зря Ваш фильм пестрит вырезками из наших статей — но отчего-то без ссылок на первоисточники).
Елена ханум, спасибо за не ангажированный ответ.
С учетом того, что Вы с Олегом (несмотря на сильное давление на вас со стороны ура-патриотов) не скрыли роль финно-угорских народов в этногенезе северных русских, я думаю, что могу Вас попросить Вас пересмотреть Вашу ныне тиражируемую теорию о генофонда тюрков, как — «доминирование элит».
Это, что за «элита», которая смогла распространить практически единый язык общетюркский язык (с учетом базисной лексики) на 10 тыс. километров в разной этнической, антропологической, религиозной среде. С учетом того, что кроме тюрков скотоводов всегда были и тюрки земледельцы (чуваши, уйгуры).
Для того, чтобы Вы поменяли Вашу точку зрения, я думаю, что достаточно будет посмотреть ещё три моих видеофильма (например, при дальнем перелете Москва-Нью-Йорк):
https://www.youtube.com/watch?v=NhbsfpGWvgU
https://www.youtube.com/watch?v=4GmspWBMNlM
https://www.youtube.com/watch?v=SlGYpkC62Mg
Заранее благодарю. С уважением.
Да мы ничего никогда не скрываем — ни про славян, ни про финно-угров, ни про алан, ни про тюрков.
А «доминирование элиты» — не наша теория. Вы нам льстите.
НАДЕЖДА МАРКИНА Этногенез и синтез наук.
Полит.ру.
Елена Балановская: Да. Конечно, в тех регионах, где эволюция языка и рост популяционной системы шли параллельно, мы наблюдаем их удивительную синхронность. И здесь лингвистика и генетика могут взаимно проверять друг друга – и любые расхождения дают пищу для размышления и толчок новым исследованиям.
Такая синхронность уже показана для народов северокавказской семьи языков – показано сходство не только родословных деревьев генофондов и языков, но даже датировок генетических и лингвистических.
У тюркских народов иная историческая модель связи генофонда и языка – в генетике она называется модель «доминирования элиты», когда смена языка связана с заменой лишь небольшой «социально верхней» части генофонда.
http://polit.ru/article/2015/03/07/klejn_balanovskaya_1/
Я все же надеялась, что Вы сужденья черпаете не из «забытых газет времен … и покоренья Крыма».
Неужели в полит.ру можно приводить массив научных ссылок на историю термина?
А здесь имела ввиду статью [Yunusbayev et al., 2015], конечно.
Дай бог, здоровья Юнусбаеву.
В последнее время. Когда я вижу или слышу некоторые тюркские фамилии, то волей- неволей вспоминаю героев знаменитого романа покойного Чингиза Айтматова. Поэтому для меня лучше почаще слышать такие фамилии как: Балаганская, Балановская, Тур, Чикишева, Волков, Степанов, Казарницкий и др, чем, фамилии якобы моих дальних соплеменниеов.
С уважением.
Современные потомки скифов.
https://independent.academia.edu/ГахраманГумбатов
Добрый день!
Уважаемая администрация , а нельзя ли забанить представителя «Великого Турана» и ему подобных? У них же , что бы им не объясняли, всегда будет свое дилетантское мнение. Просто обидно , что профессионалы тратят свое время на подобные дискуссии…
Елена Владимировна, попутно вопрос — ожидаются какие-либо публикации, касающиеся Самарской, Хвалынской, Средний Стог и других культур раннего неолита Ю-В Европы?
Уважаемый Борис!
Мы забанили уже многих. Но обычно все же даем возможность высказаться человеку, если он не переходит границ. Чтобы не тратить время, можно просто не отвечать.
Елена Владимировна сейчас в отпуске, так что вряд ли сможет Вам ответить в ближайшее время. Возможно, она будет смотреть почту и увидит Ваш вопрос. Но если ответа не будет, то по этой причине.
Всего доброго,
Надежда Маркина
модератор
Спасибо Надежда!
Елене Владимировне удачно отдохнуть)
Просьба. Вы когда картинки размещаете, делайте так чтобы они на самом деле увеличивались, а не просто открывались как у вас. Не видно процентов на шкале на картах, смотреть вообще неудобно.
Уважаемый Иван, это вряд ли возможно технически. Картинка открывается до максимального ее разрешения.