
Одной из пограничных областей, в которой особенно ярко проявляются контакты между науками, является анализ древней ДНК, потому что тут работают совместно генетики, археологи, палеоантропологи, а зачастую привлекаются и лингвисты. Поэтому первая часть доклада будет посвящена древней ДНК, во многих исследованиях которой мне довелось участвовать. Анализ современных генофондов, хотя и находится в ведении только одной науки – популяционной генетики – но и эти результаты активно обсуждаются самыми разными специалистами, и это вторая часть доклада. Наконец, сотрудничество генетиков и этнографов – один из ярких примеров междисциплинарного подхода, но оно иногда наталкивается на недопонимание того, в чем совпадают, и в чем различаются этнос (объект этнологов) и популяция (объект генетиков).

Часть I Древняя ДНК
Идея изучения древней ДНК очень простая – взять древнюю косточку и сравнить ее ДНК с ДНК из всех современных популяций. Но технически это сложно: в выдыхаемом нами воздухе содержится больше ДНК, чем во всей этой косточке. И для того, чтобы изучать не свою ДНК, а ту, которая содержалась в этой косточке, нужно надеть скафандры и 90% времени тратить на поддержание стерильности помещения. В мире есть некоторое количество лабораторий, которые занимаются этим достаточно надежно.

Один из вопросов, который привлекает больше всего внимания, это неолитизация Европы. У исследователей сложилась такая точка зрения, что генофонд Европы сформировался в ходе двух событий– во-первых, когда люди туда попали впервые, и, во-вторых, когда туда пришли земледельцы. Ведь появление земледелия (производящего хозяйства) привело к многократному увеличению численности, поэтому вклад этой миграции многократно важнее, чем вклад последующих. И сейчас господствует точка зрения, что генофонд Европы сформировался под влиянием миграции населения с Ближнего Востока. Согласно теории демической диффузии, неолитическая популяция, расселяясь, ассимилирует автохтонное население, формируя градиент в генофонде. И этот градиент подтверждается данными последних исследований. Земледельцы двигались с Ближнего Востока, постепенно смешиваясь с местным населением, и формируя генофонд Европы. Однако данные по митохондриальной ДНК указывают на более древние датировки формирования европейского генофонда, чем неолит, что не согласуется с такой концепцией.
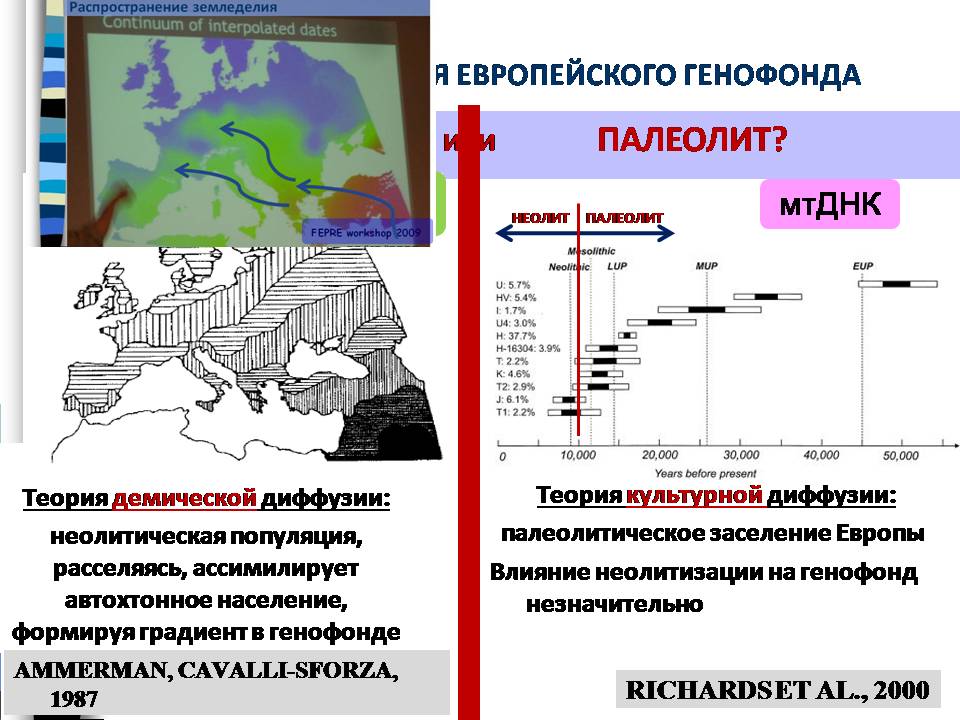
Один из способов разрешить это противоречие, это непосредственно посмотреть генофонд первых неолитических земледельцев Европы и разобраться, местные ли это уроженцы, которые восприняли технологию земледелия, или это мигранты издалека.

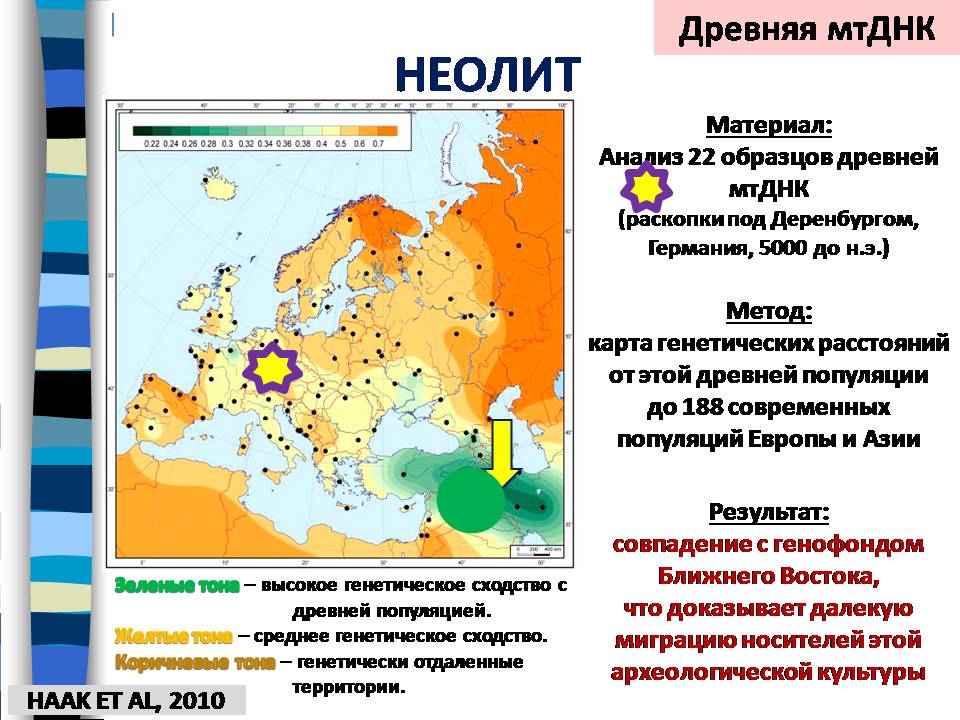
На карте ниже показаны результаты сравнения 22 образцов древней мтДНК, найденных на территории Германии, с 188 современными популяциями Европы и Азии. Методом генетических расстояний показано сходство древней мтДНК с генофондом Ближнего Востока. Так что не приходится сомневаться, что была миграция древнего ближневосточного населения, земледельцев. По-видимому, она была не слишком многочисленна, и генетически растворилась в местном населении.
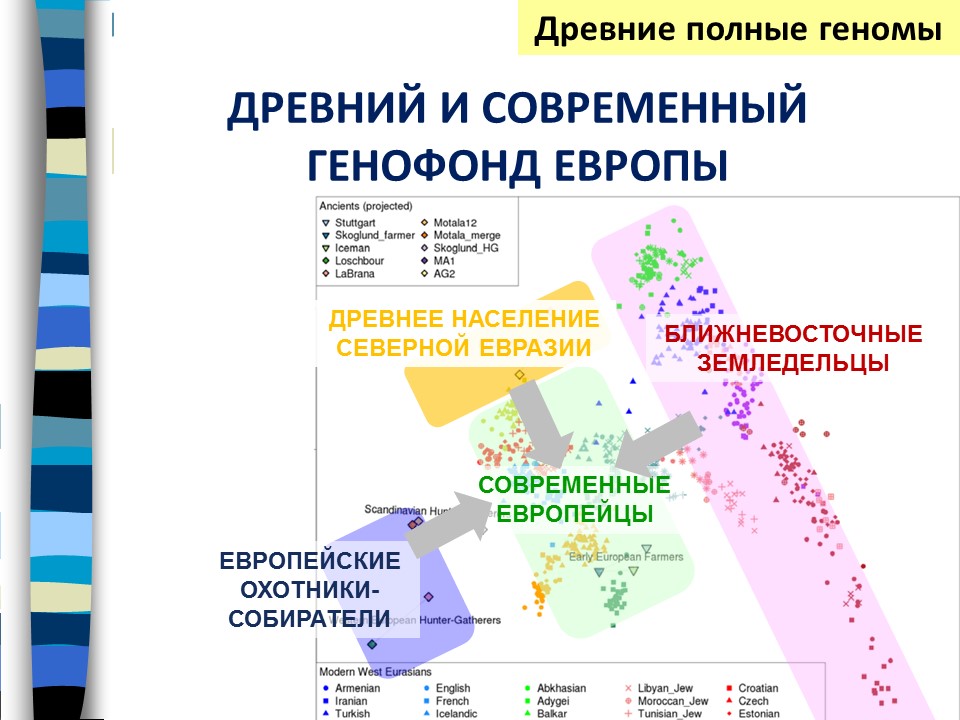
С появлением возможности исследования полных геномов, в том числе древних ДНК это представление получило наглядное графическое подтверждение. На этом рисунке видно, где находится генофонд населения Ближнего Востока, где – геном европейских охотников-собирателей, и что геном современных европейцев находится посередине между ними. Так что выявляется смешение этих групп населения. Но выявляется и нечто третье – это древний евразийский компонент, который тоже вносит свой вклад в генофонд Европы. Статья о взаимодействии этих трех компонентов была опубликована в Nature.
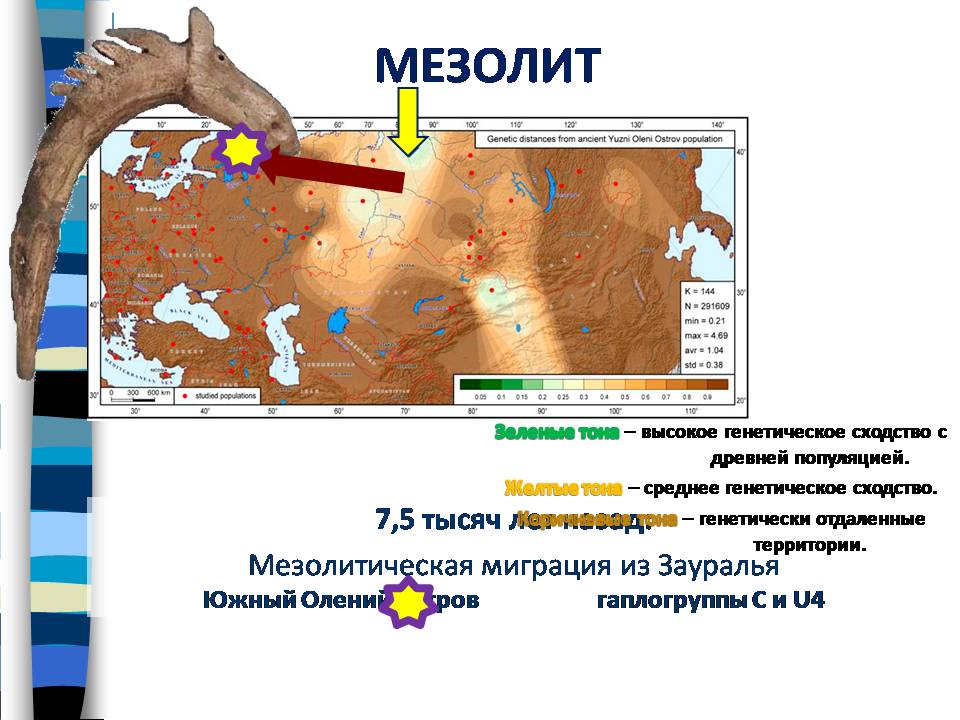
Если обратиться к более древней эпохе – к мезолиту, то была изучена популяция мезолитических охотников-собирателей Оленеостровского могильника на Южном Оленьем Острове в Онежском озере. А зона генетически сходных с ними популяций, как показывает карта, оказалась в Сибири. И это указывает на миграцию, произошедшую в мезолите из Сибири на территорию Европы.
Была изучена жившая примерно в том же регионе, но на 3 тысячи лет позже, популяция с Большого Оленьего острова, которая тоже обнаружила сходство с популяциями Сибири. Это доказывает, что миграции из Сибири были множественными. Интересно, что современное население этих мест не имеет ничего общего с той древней популяцией. То есть, та древняя популяция не оставила ныне живущих потомков, а значит, установить эту миграцию по современному населению мы бы никогда не смогли.

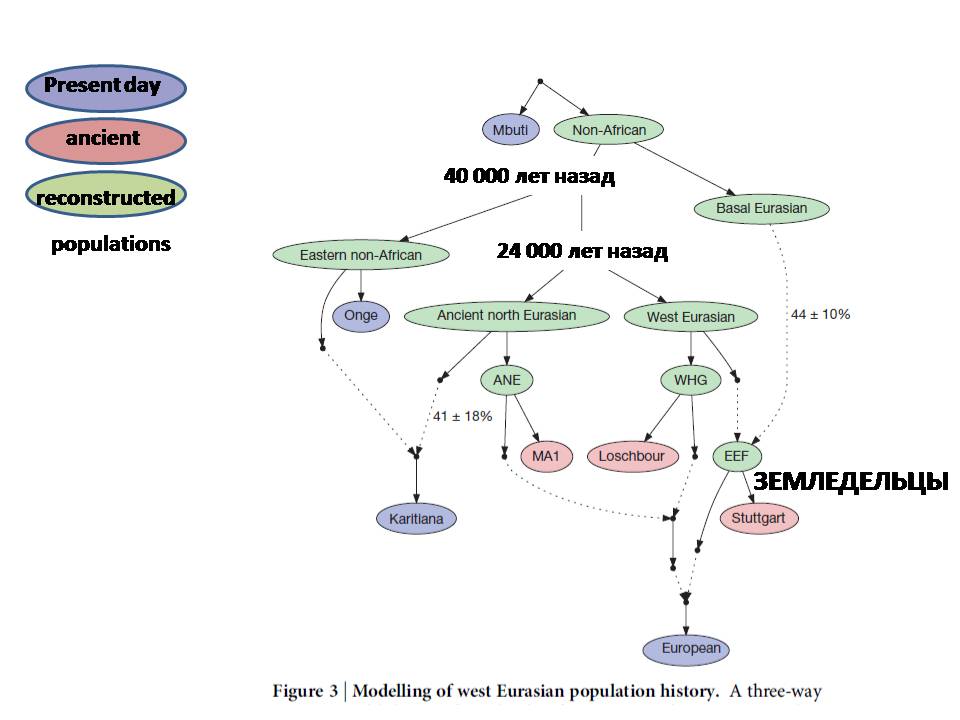
Приведу одну из самых недавних глобальных реконструкций, в которую включены данные и по древней, и по современной ДНК, эта некая схема ветвления человечества. Тут есть разделение на африканцев и неафриканцев, потом отделение некой загадочной ветви «базальных европейцев», последующее разделение населения Северной Евразии и Западной Евразии и объединение некоторых из этих потоков. Здесь важно то, что сейчас мы имеем возможность не реконструировать некоторые отдельные направления миграции, а осознавать всю сложность популяционной системы – как разделение, так и смешение.
Часть II Современные генофонды
Перейдем от древней ДНК к современной. Наука геногеография – в широком смысле это наука, изучающая генофонды. В узком понимании генетическая география — это карты. Справедливость этого в том, что карта действительно является сердцевиной всей геногеографии. Пространство — это гораздо более важное измерение для изучения генофонда, чем время. В этом году геногеография могла бы праздновать свой столетний юбилей, потому что впервые идея о том, что частоты генов могут быть у людей разными, что на одной территории гены встречаются с одной частотой, а на другой территории могут быть с совсем другой частотой – возникла из практики Первой мировой войны.

Когда стало необходимым массовое переливание крови, выяснилось, что для переливания крови русским, сербам, французам, неграм, индусам и т.д. нужно запасаться разными объемами одних и тех же групп крови. С этого и начались исследования разнообразия человечества по генетическим маркерам. За сотню лет опубликованы десятки тысяч статей по разным типам генетических маркеров. Сначала использовались классические иммунологические маркеры, потом перешли к классическим биохимическим, потом началась эра ДНК. Существует очень много типов генетических маркеров, но сейчас используются три основных типа: Y-хромосома, митохондриальная ДНК и полногеномные маркеры. По времени использование этих пяти типов маркеров почти последовательно сменяет друг друга. Например, сейчас стремительно растут и будут расти исследования полногеномных маркеров.
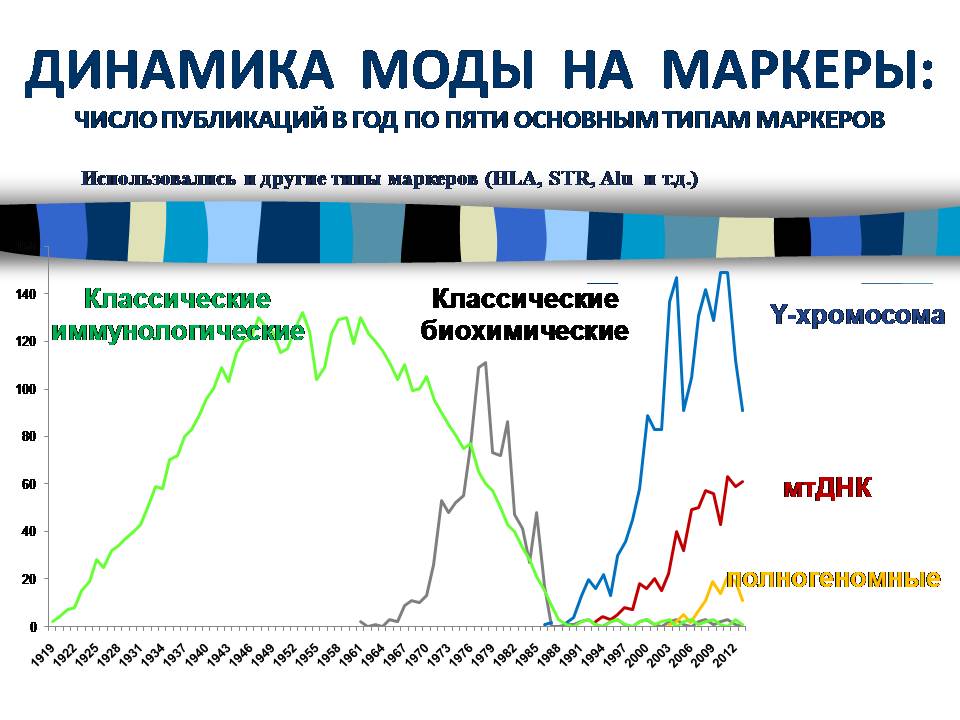
Я продемонстрирую основные исследования за последние полвека по этим всем маркерам, чтобы показать, как стабильность, то есть постоянно выявляемые одни и те же результаты, так и динамику.
Переход от старых маркеров к ДНК стал неким испытанием, потому что можно было предполагать, что ДНК даст нечто совсем другое. Но на самом деле результаты по старым маркерам стали прогнозом для новых. Этот принцип с тех пор неоднократно подтвердился.

Это позволяет нам ввести полисистемный подход, то есть когда у нас присутствуют разные типы маркеров, разные признаки, мы можем изучить какие-то популяции по разным маркерам и сравнить результаты. Если они окажутся разными, это либо ошибки, либо особенности восприятия разных маркеров, которые указывают на объективные черты генофонда, но которые по-разному преломляются в призмах разных маркеров.

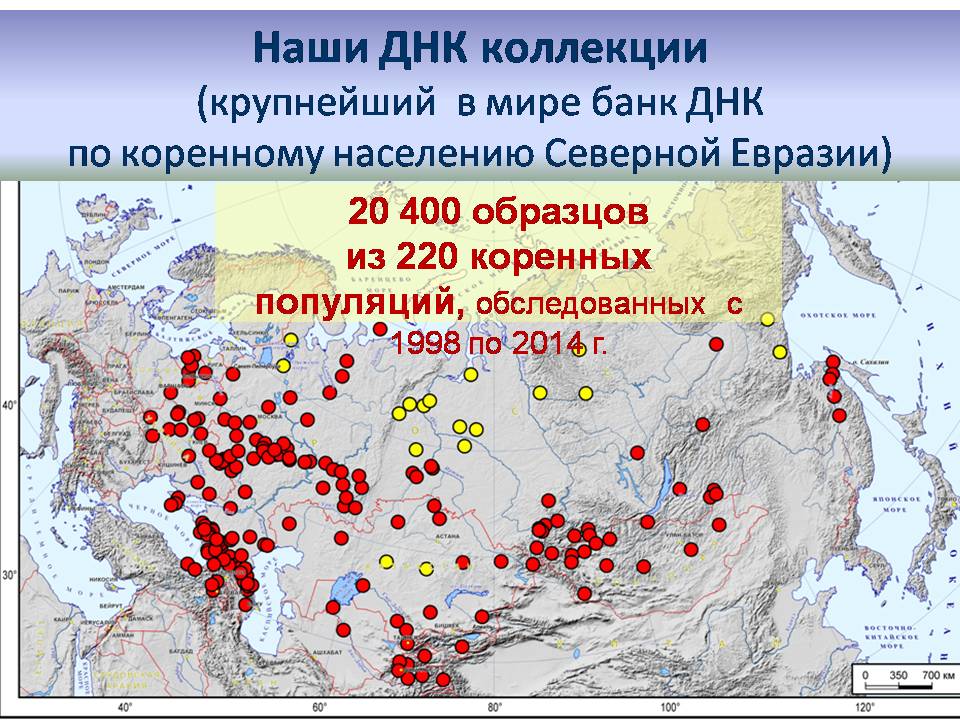
На следующей карте показаны популяции, обследованные в наших экспедициях. Наши ДНК-коллекции – это крупнейший в мире банк ДНК коренного населения для Северной Евразии. С 1998 по 2014 год мы собрали 20400 образцов из 220 коренных популяций. Но в базы данных, которые м ы создаем и которые используются для всех анализов, включены не только наши данные, но и почти все то, что публикуется в мире.
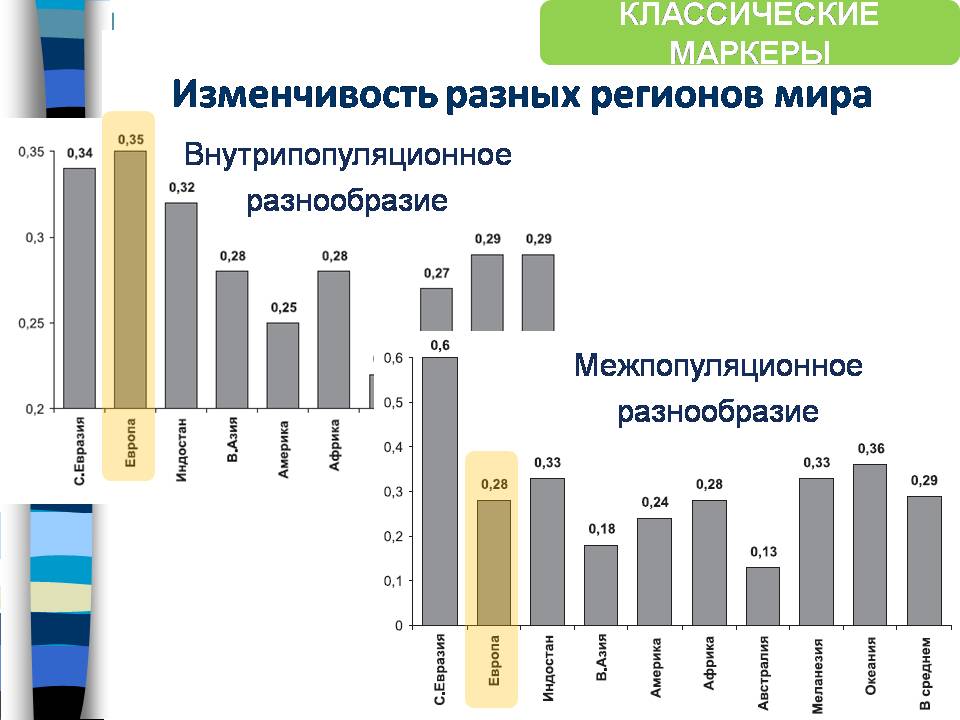
По классическим маркерам можно сравнить внутрипопуляционное и межпопуляционное разнообразие. Например, если посмотреть на Европу, то видно, насколько велики внутрипопуляционные различия и низки различия между популяциями.
Другим методом получены карты главных компонент, на которых переход цвета соответствует важнейшему направлению генетической изменчивости.
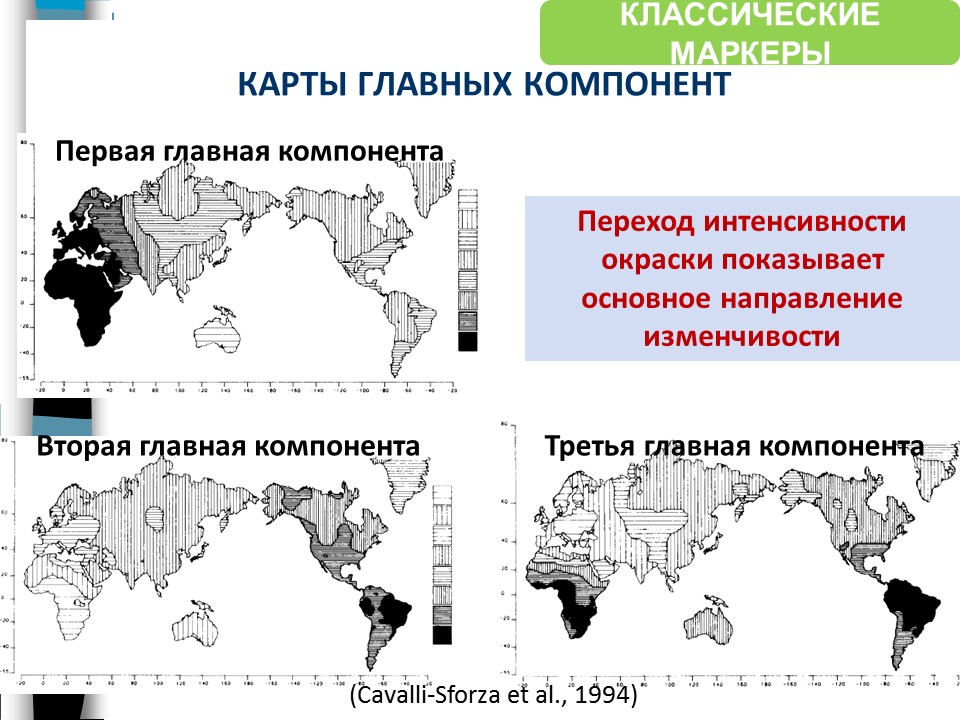
Главная компонента не одна – их столько, сколько исходных признаков, но важны только первые две-три из них. Так, по первой главной компоненте мы видим кардинальные различия между населением Африки и населением Австралии, а все остальное – в промежутке. Вторая главная компонента показывает основные различия между населением Африки и населением Южной Америки. Третья – изменчивость между западом и востоком Евразии.
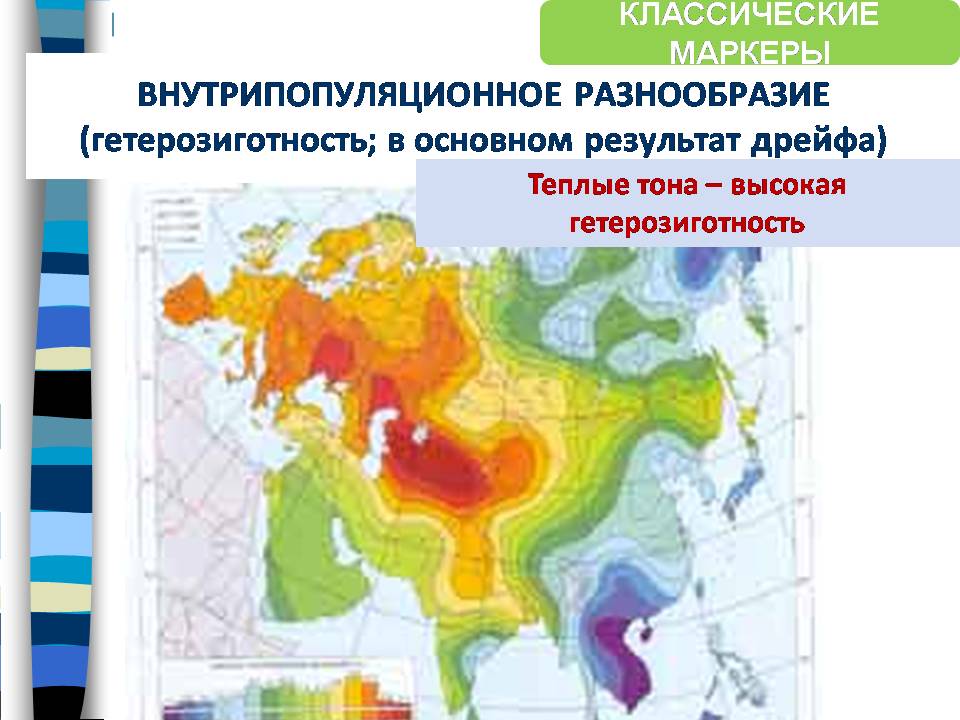
На следующей карте изображено внутрипопуляционное разнообразие, видно, что оно наиболее высоко в западных и южных областях Евразии.

Эти данные (на рисунке выше) получены зарубежной школой геногеографии, а независимо работала отечественная школа Рычкова (рисунок ниже), и результат они получили тот же самый.
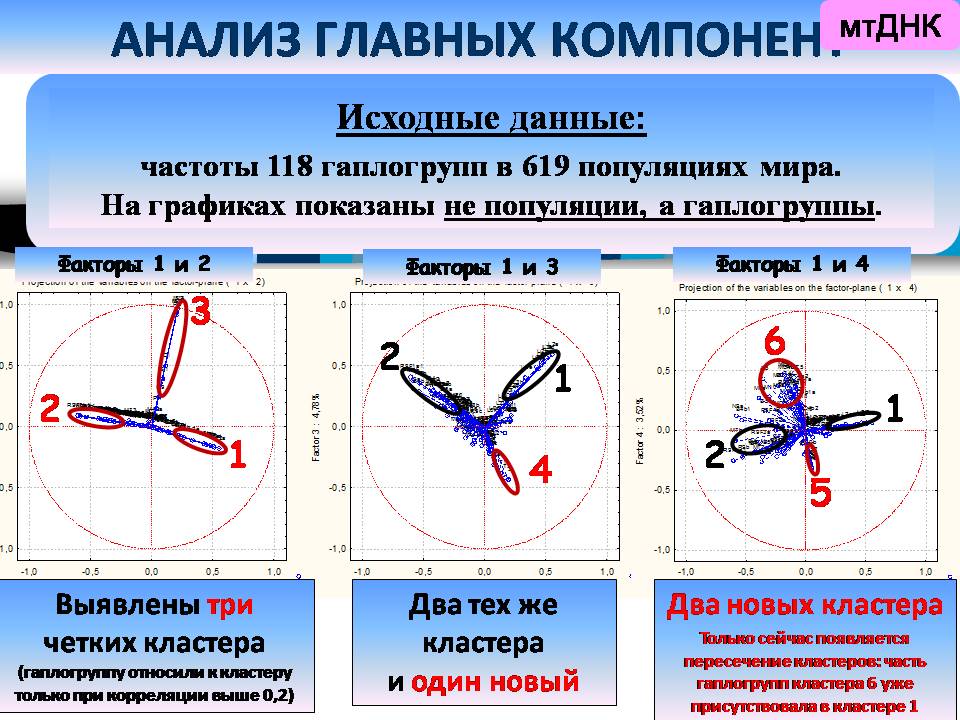
По мтДНК были проанализированы более сотни гаплогрупп в более чем 600 популяциях мира. Мы посмотрели корреляции разных признаков друг с другом. Видно, что признаки образуют неожиданное четкие кластеры по их пространственной корреляции друг с другом, и всего выявлено шесть таких главных кластеров. Сто восемнадцать признаков сходятся всего к шести основным паттернам изменчивости! Рассмотрим эти паттерны.
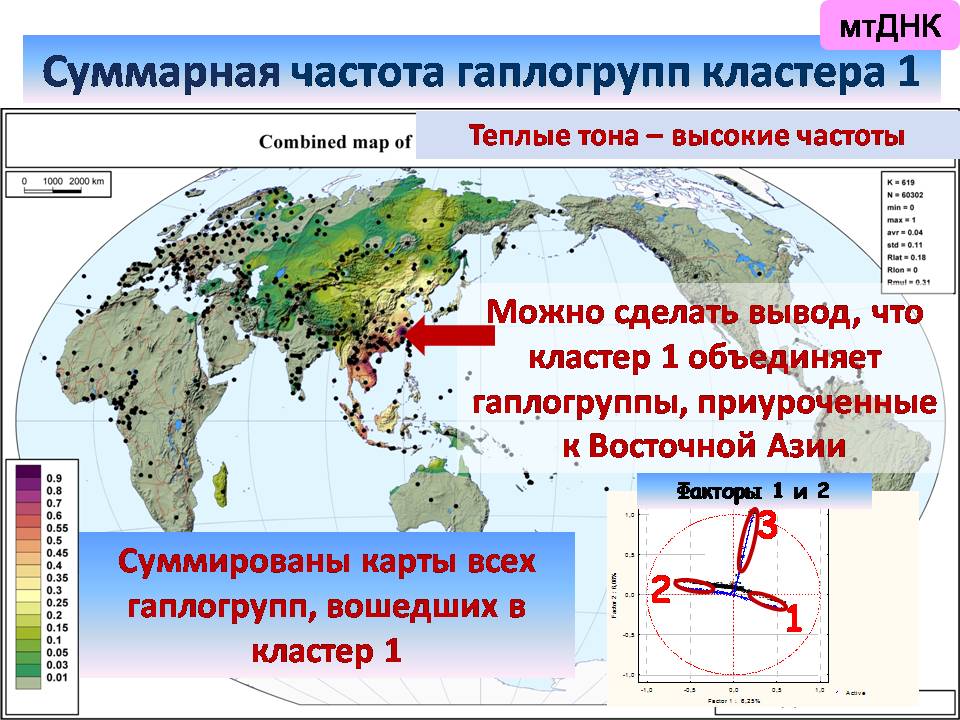
Если суммировать частоты гаплогрупп, входящих в разные кластеры, то видно, что первый, самый многочисленный кластер приурочен к Восточной Азии,
второй кластер – однозначно африканский,
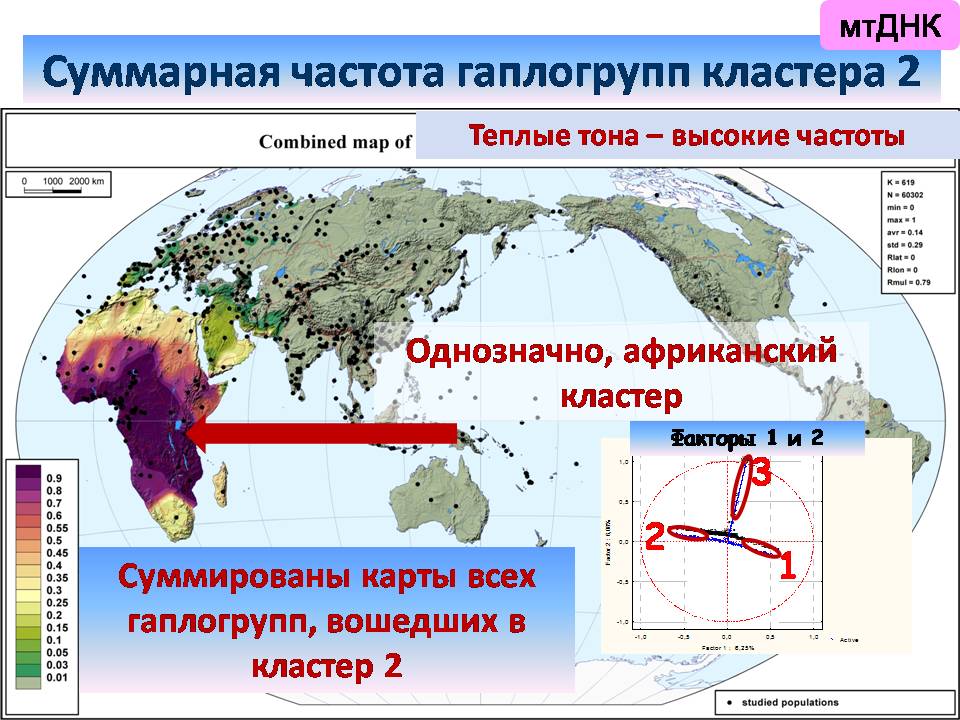
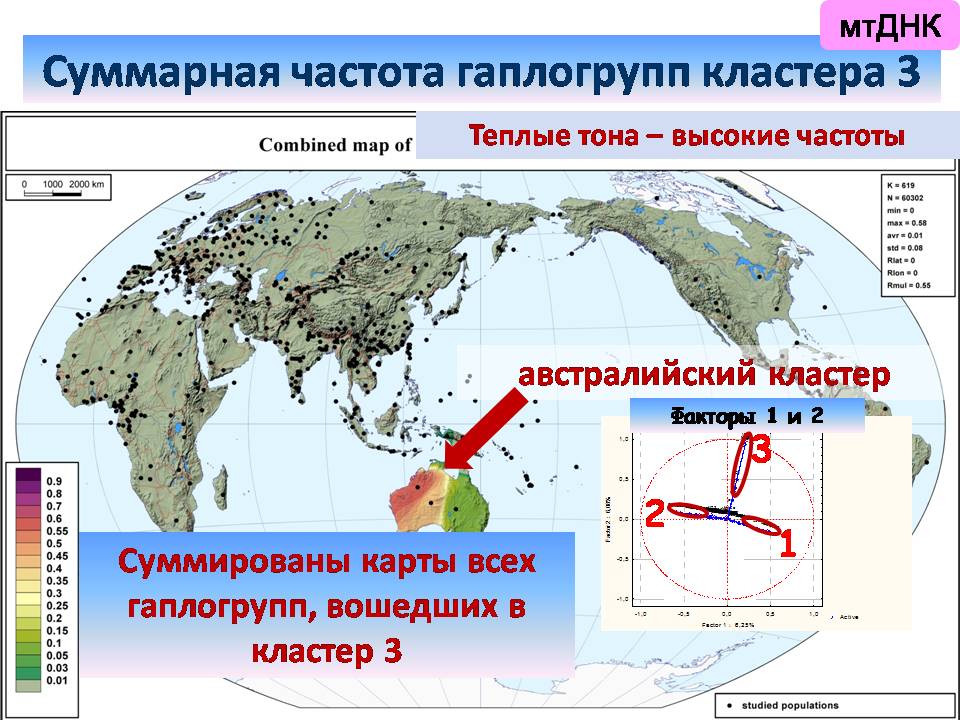
а третий оказывается приурочен к Австралии.
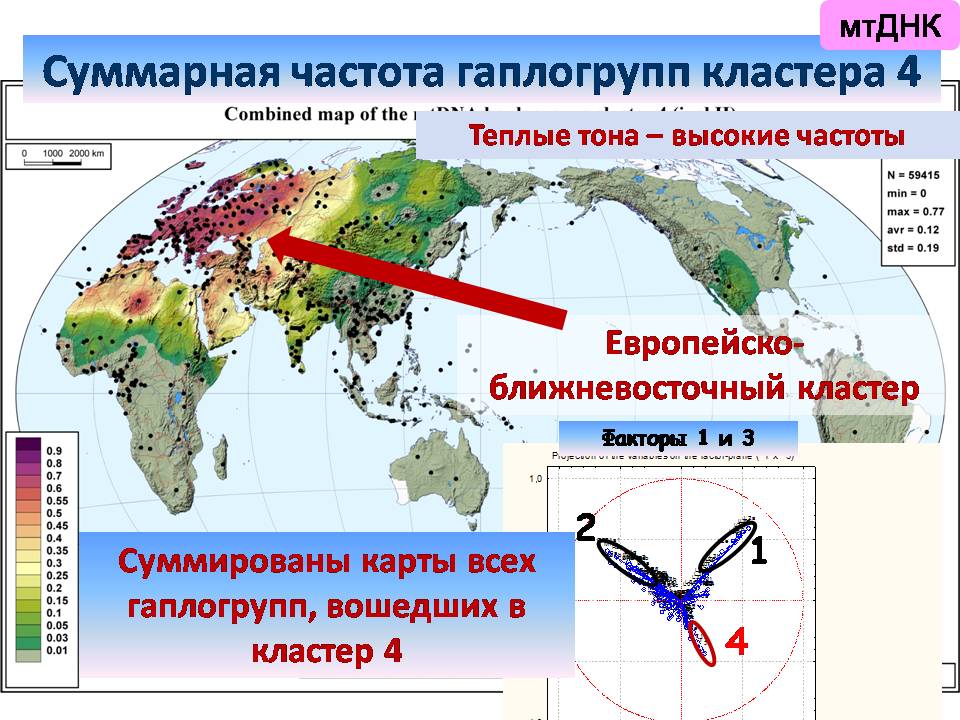
Затем у нас выделяется западноевразийский кластер,
и в отдельный кластер выделилась Сибирь.
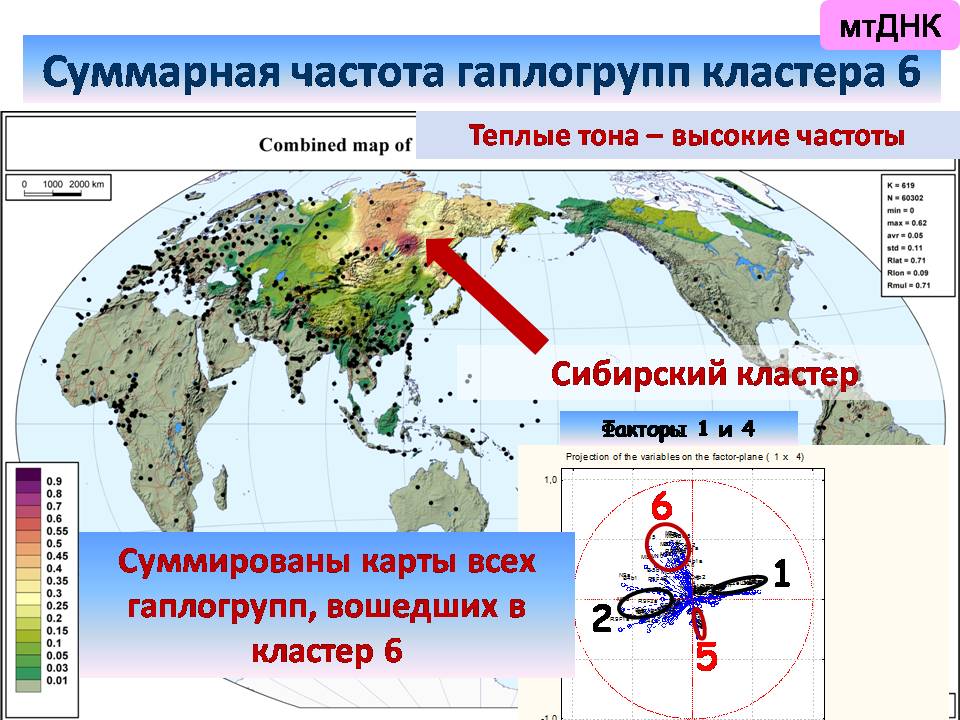
Это означает, что по мтДНК северные и южные монголоиды (кластеры 1 и 6) повышаются в ранге до вполне самостоятельных континуумов, сравнимых с африканским и австралийским.
Итак, по мтДНК выявлены те же самые геногеографические провинции мира, которые выявлялись и экстремумами главных компонент.
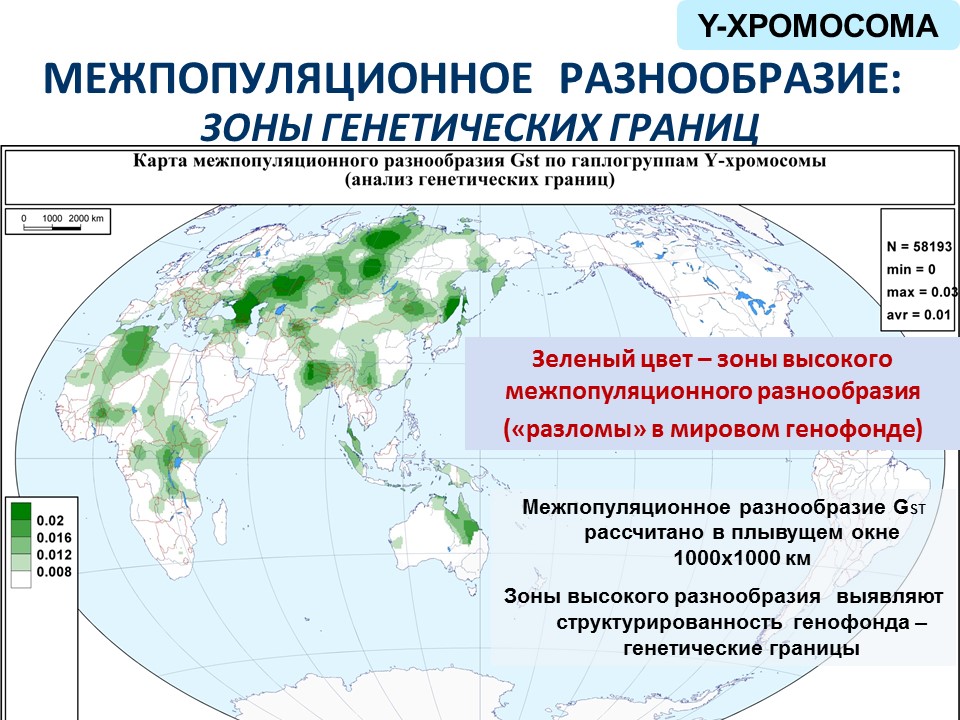
Говоря про Y-хромосому, выяснилось, что если проанализировать данные теми же методами, то мы получим похожие результаты. И поскольку времени недостаточно, для Y-хромосомы я не буду показывать континуумы гаплогрупп и внутрипопуляционное разнообразие, покажу анализ межпопуляционного разнообразия. Оно может быть большим на стыках контрастных, различающихся генофондов – так же, как на стыках двух материковых плит возникают горы. Примерно то же самое происходит и в генофондах. И выделяются зоны высокого популяционного разнообразия – «разломы» в мировом генофонде.
Например, одна из таких зон – это Гималаи, по разные стороны Гималаев очень разное население. А основной разлом проходит наискосок от Кавказа до Средней Сибири и делит Евразию пополам.
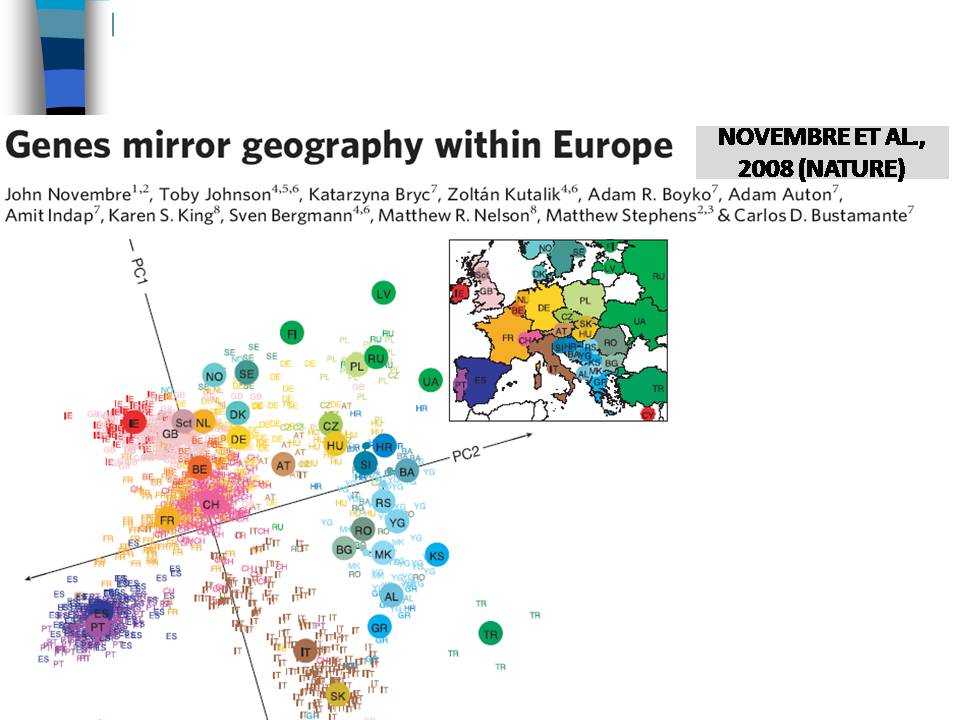
По полногеномным маркерам покажу только один результат – если в генетическом пространстве разместить отдельных индивидов, то люди, живущие в определенных регионах, хорошо кластеризуются друг с другом.
Часть III Генетики и этнологи
Из всех этих и многих других сведений о структуре генофонда какую информацию можно извлечь об истории популяций и происхождении народов? Обратите внимание, что до сих пор я ни разу не упомянул этносы. Привязка данных геногеографии к этносам является далеко не самым главным в изучении генофонда человечества. Но именно это вызывает наибольшее количество вопросов и наибольшую степень непонимания. Геногеография все время находится между Сциллой и Харибдой. Пучина Харибды – утверждение о том, что не существует никакой связи между генетикой и этносом. Но ужасы Сциллы еще хуже – она утверждает 100% связь между генетикой и этносом. Это то, к чему склоняется ДНК-генеалогия.

Как же все-таки пройти между Сциллой и Харибдой? Для этого надо задуматься, что если генетика имеет дело с биологическими объектами, с популяциями, откуда там вообще возникает этнос – категория социальная? Дело в том, что популяция для популяционной генетики – это любая группа людей, которая удовлетворяет всего-навсего двум условиям. Она должна существовать более чем одно поколение, и более половины браков должно заключаться внутри нее. Любая группа людей, удовлетворяющая этим критериям, является популяцией.

Обозначим людей кружками, а заключаемые ими браки – стрелками. Тогда совокупность людей, обрисованная красной линией, является популяцией по критерию 50% браков, потому что более половины стрелочек, которые начинаются внутри этой линии, внутри нее и заканчиваются и только 4 стрелки выходят за ее пределы.
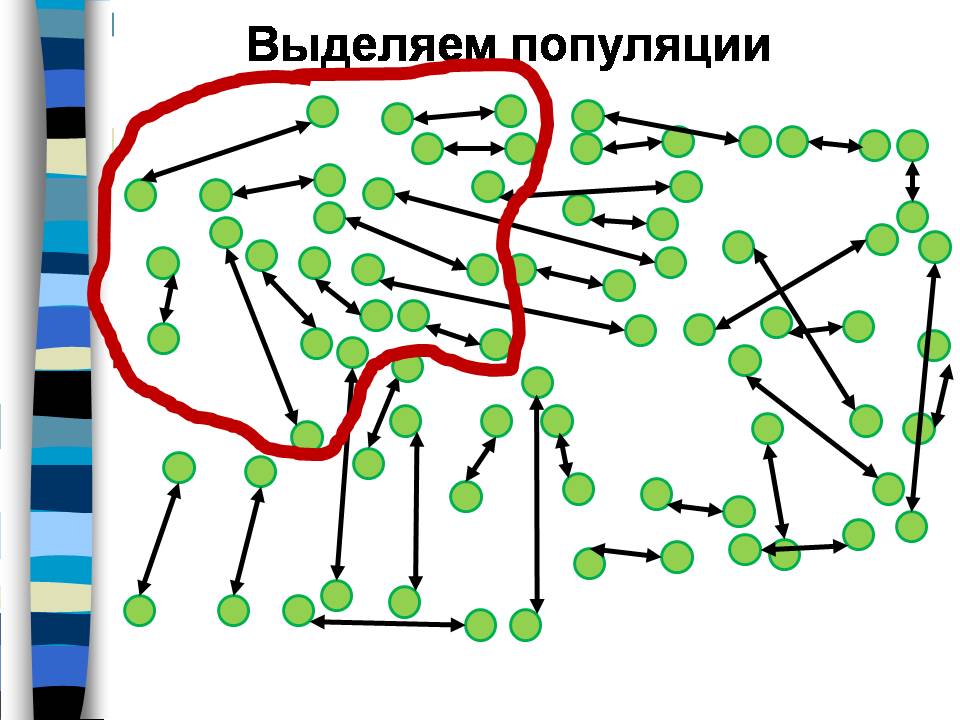
Группа людей, ограниченная фиолетовой линией, тоже является популяцией, потому что опять большинство браков заключены в ее пределах.
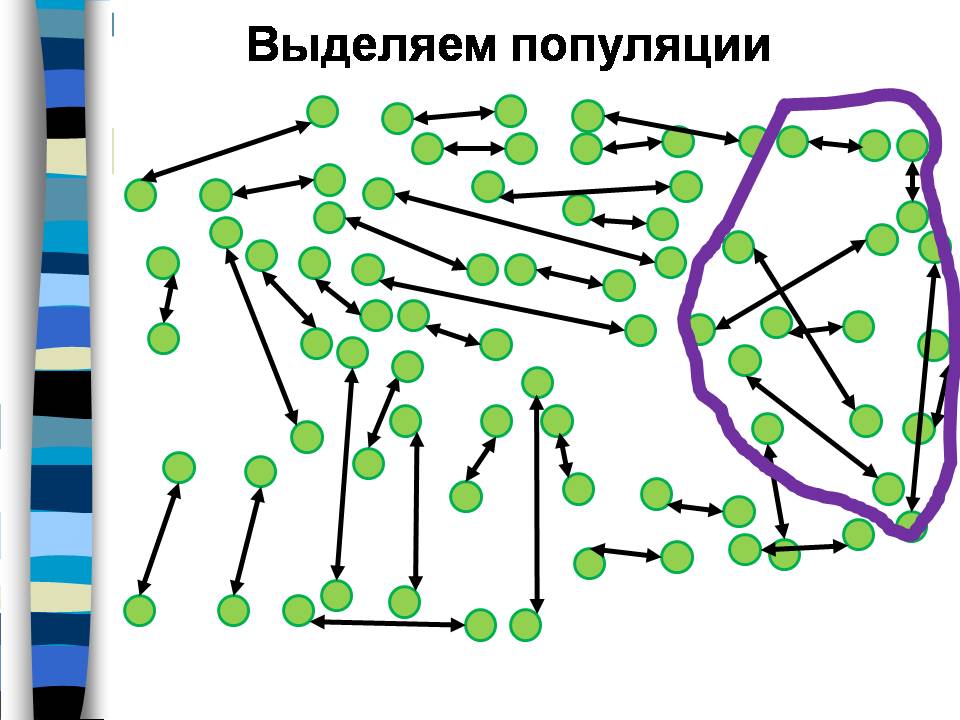
И группа людей, ограниченная желтой линией, тоже является популяцией. Она включает часть людей из первой популяции, и часть из второй. На этом примере наглядно видно, что популяции можно выделить разными способами, они могут быть вложенными друг в друга или перекрываться.
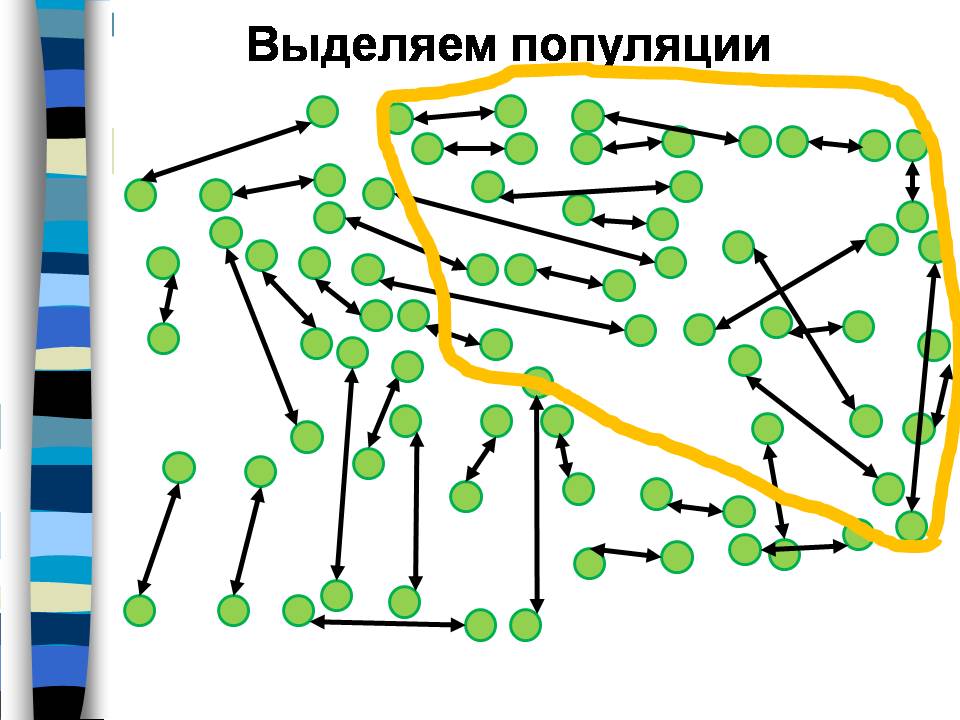
А является ли популяцией группа людей, ограниченная розовой линией? Давайте посчитаем: в этой группе 7 человек, но только двое из них заключили брак в пределах этой группы, а остальные 5 (большинство) – за ее пределами.

Когда доступны такие полные данные по структуре браков, выясняется, что, например, в Архангельской области – если взять одну деревню, то внутри нее заключается меньше 50% браков – это не популяция. Но если взять несколько соседних деревень, то уже больше 50% — это уже популяция.
А на юге – если взять весь Краснодарский край, то больше половины браков заключены с уроженцами других регионов. Значит, население всего Краснодарского края не является популяцией.

Популяции входят одна в другую, как матрешки, они перекрываются. Человечество представляет собой сложнейшую систему популяций. И поскольку полные данные по структуре браков обычно недоступны, то популяционным генетикам для выделения популяций нужны какие-то другие параметры, которые с брачной структурой скоррелированы. Поэтому любые признаки, по которым существует брачная ассортативность, являются признаками, очень полезными для популяционных генетиков.

Например, географический признак – он работает почти всегда – как правило, большинство браков заключается с уроженцами не слишком отдаленных регионов. Конфессиональный – например, популяция мормонов США – это классический объект популяционной генетики, на котором сделано много работ. Социальный – популяция людей с высшим образованием. Языковой, например, популяция русскоязычных, сословный – популяция казаков. Наконец, этнический.

Для самых разных популяций максимальная ассортативность браков происходит по принципу географического соседства и по формальному критерию этничности – той этничности, которую люди себе указывают.

Итак, для каждого человека, биологический образец которого анализируется популяционными генетиками, фиксируются как минимум два параметра: его происхождение из одного из известных регионов (из списка) и происхождение от одного из народов (тоже из списка).

Возникает вопрос, а откуда берутся эти списки и кто, вообще говоря, определяет, что такое регион и что такое народ?
Вопрос, что такое регион, очень простой — его решает правительство, а генетика принимает то, что говорит правительство. В данный момент господствует практика, что географические границы регионов жестко закреплены, заданы на географических картах и считаются неотъемлемыми свойствами регионов. Генетика это принимает. Если же завтра выйдет закон, по которому границы регионов являются воображаемыми, конструируются самими жителями регионов и легко изменчивы, генетика примет и это. Если когда-то регионы будут вообще отменены, а место рождения будет записываться, как GPS-координаты, генетика примет и это. Мы будем работать с такими данными и выделять популяции по принципу близости GPS-координат.

А вопрос, что такое этнос, решает этнология. Генетика принимает то, что говорит этнология. До недавнего времени существовала единственная концепция эссенциализма, которая воспринимает принадлежность к определенному этносу как изначальное и неотъемлемое свойство людей. И мы с этим работаем. Недавно, наряду с эссенциализмом, появилась и концепция конструктивизма, которая рассматривает этнос как воображаемую конструкцию, создаваемую самими людьми и легко изменчивую. И мы это тоже принимаем, и работает с таким пониманием этноса. Если в будущем в этнологии возникнут иные концепции, генетика примет и их.

Генетику интересует только одно – по какому признаку ассортативности заключаются браки. Но тут возникает вопрос о стабильности признака в поколениях, потому что если признак нестабилен, то он становится неинформативен для выделения популяций. И вот более 20 лет назад на одном из докладов мы представили пример из нашей экспедиционной практики на Сахалине, когда человек, оба родители которого – нивхи, объявил, что он норвежец.

Вопрос тут число количественный. Если место рождения для большинства членов популяции станет изменяться от поколения к поколению, генетика перестанет учитывать место рождения как информативный для генетики показатель. Но пока более 50% детей характеризуются тем же регионом рождения, что и их родители, место рождения учитывается и помогает нам работать.

Точно так же, если параметр этничность для большинства членов популяции станет изменяться от поколения к поколению, генетика не станет его учитывать.

Если взять группу людей одинаковой этничности, то есть вероятность, что среди их потомков часть будет причислять себя к другому этносу. Но если таких потомков меньше 50%, то признак информативен, этот этнос – популяция. А вот если этничность изменится у более 50% детей, значит, этот этнос не популяция. И тогда признак этничности для генетики не работает.
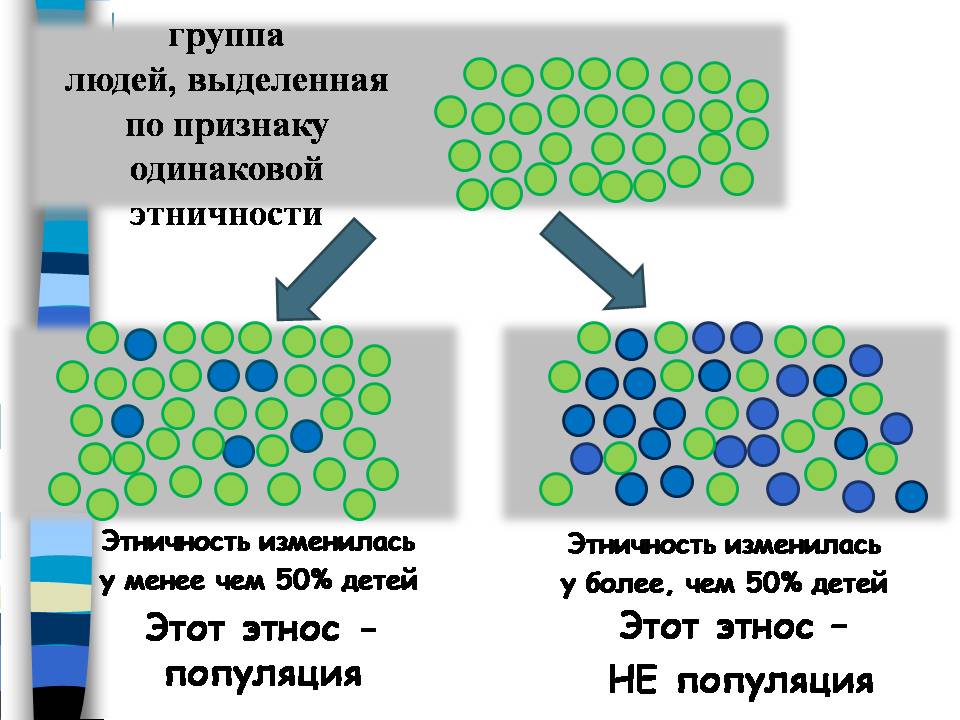
Реальные данные такие – в ходе наших исследований мы провели анкетирование более 20 тысяч человек из 55 этнических групп. И ни для одной этнической группы смена этнической идентичности за одно поколение не достигала не только 50%, но даже и 5%. Поэтому эти два параметра – этническая самоидентификация и место рождения, пока отражают объективную реальность и остаются важнейшими критериями выделения популяций.

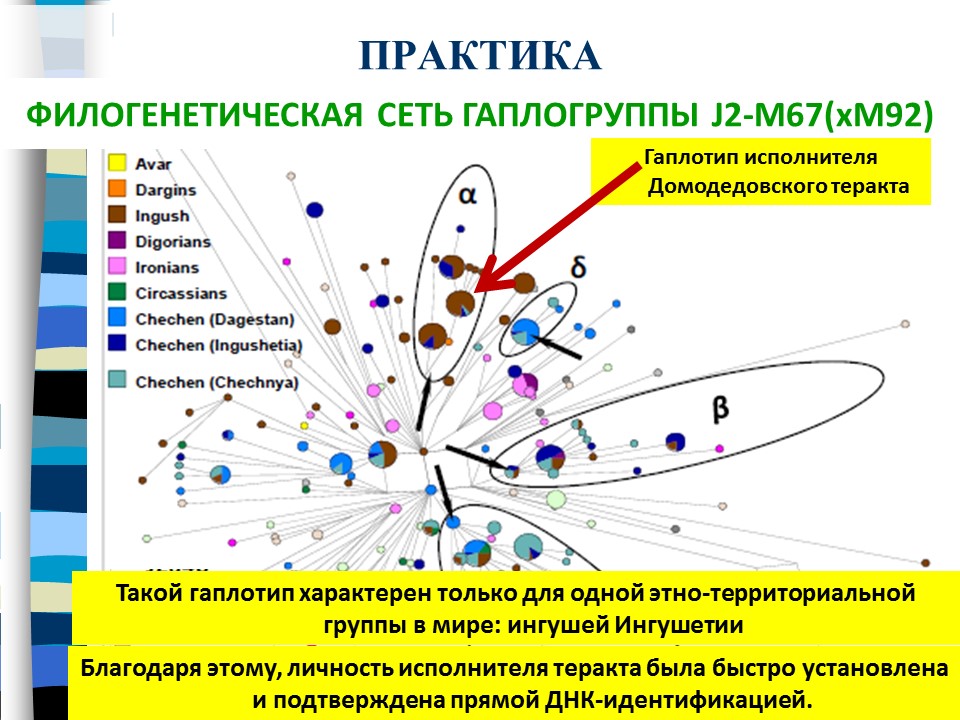
Практическая эффективность такого подхода была выявлена в том случае, когда по Y-хромосомному гаплотипу исполнителя домодедовского теракта удалось определить место его рождения. Этот гаплотип точно совпал с группой, которая была встречена только у одной группы в мире – ингушей Ингушетии. Благодаря этому личность исполнителя была быстро установлена и подтверждена прямой ДНК-идентификацией.
Это не единичный случай – Следственный комитет обращается к нам ежемесячно.

Сотрудничество с этнографами для генетиков очень важно, и очень многие этнографы отвечают нам взаимностью.




Среди нашей большой команды разнообразны места рождения.

И разнообразна этническая принадлежность.

А вот браков внутри нашей команды всего 4%. Поэтому наш коллектив не является популяцией. Мы являемся частью общей популяции научного сообщества и открыты для взаимодействия с другими коллективами.

видеозапись доклада https://www.youtube.com/watch?v=YmcM2MifMeU
Добавить комментарий
Для отправки комментария вам необходимо авторизоваться.